„das Hohlraumsystem der Tentakelkrone ungleich einfacher“ sein würde als nach meinen Auseinandersetzungen.
Das Fadengewirr innerhalb des Kanals bei Cristatella halte ich noch heute für die Schwänze
der zusammengetriebenen Spermatozoen, die sich auch hier und da in den übrigen Theilen der Lophophorhöhle
nachweisen lassen. Neben den Samenfäden finden sich die Restkörper der Spermatoblasten. Nur
an Thieren, welche notorisch geschlechtsreif waren, habe ich derartige Ansammlungen aufgefunden.
Später zeigen sich an ihrer Stelle Conglomerate von degenirten Gewebstheilen, deren zellige Natur mir
niemals zweifelhaft war. Von Harneoncrementen sah ich keine Spur.
Da, wo Cori seine Oeffnung beschreibt, bin auch ich allerdings über eine solche im Zweifel gewesen,
nicht über eine Mündung nach aussen, sondern über eine Verbindung mit der Epistomhöhle. Die Erweiterung
des Kanals reicht nämlich etwas unter die Vereinigungsstelle der paarigen Aeste hinab — Coris Ausführungsgang
—, und es schien mir möglich, dass hier, zwischen den Aesten hindurch, eine feine Oeffnung nach
der Epistomhöhle führen könnte. Ich habe sie jedoch niemals deutlich gesehen, und in Fällen, wo der
Gabelkanal weit aufgetrieben war, konnte ich an zuverlässigen Medianschnitten hier nur eine sehr zarte
doppelte Mesodermlage nachweisen. Für ein Excretionsorgan wäre mit einer solchen Oeffnung natürlich
nichts gewonnen gewesen.
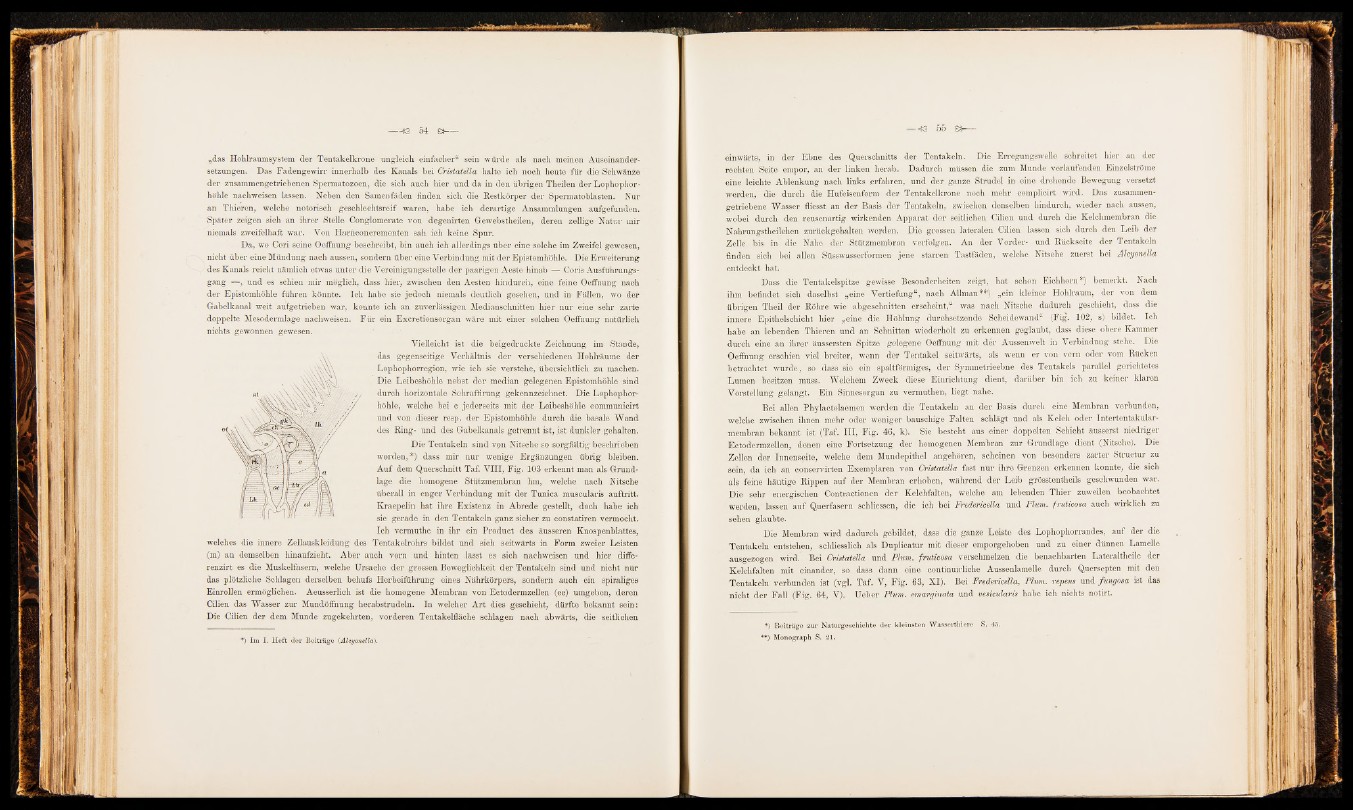
Vielleicht ist die beigedruckte Zeichnung im Stande,
das gegenseitige Verhältnis der verschiedenen Hohlräume der
Lophophorregion, wie ich sie verstehe, übersichtlich zu machen.
Die Leibeshöhle nebst der median gelegenen Epistomhöhle sind
durch horizontale Schraffirung gekennzeichnet. Die Lophophorhöhle,
welche bei c jederseits mit der Leibeshöhle communicirt
und von dieser resp. der Epistomhöhle durch die basale Wand
des Ring- und des Gabelkanals getrennt ist, ist dunkler gehalten.
Die Tentakeln sind von Nitsche so sorgfältig beschrieben
worden, *) dass mir nur wenige Ergänzungen übrig bleiben.
Auf dem Querschnitt Taf. VIII, Fig. 103 erkennt man als Grundlage
die homogene Stützmembran hm, welche nach Nitsche
überall in enger Verbindung mit der Tunica muscularis auftritt.
Kraepelin hat ihre Existenz in Abrede gestellt, doch habe ich
sie gerade in den Tentakeln ganz sicher zu constatiren vermocht.
Ich vermuthe in ihr ein Product des äusseren Knospenblattes,
welches die innere Zellauskleidung des Tentakelrohrs bildet und sich seitwärts in Form zweier Leisten
(m) an demselben hinaufzieht. Aber auch vom und hinten lässt es sich nachweisen und hier differenzirt
es die Muskelfasern, welche Ursache der grossen Beweglichkeit der Tentakeln sind und nicht nur
das plötzliche Schlagen derselben behufs Herbeiführung eines Nährkörpers, sondern auch ein spiraliges
Einrollen ermöglichen. Aeusserlich ist die homogene Membran von Ectodermzellen (ec) umgeben, deren
Cilien das Wasser zur Mundöffhung herabstrudeln. In welcher Art dies geschieht, dürfte bekannt sein:
Die Cilien der dem Munde zugekehrten, vorderen Tentakelfläche schlagen nach abwärts, die seitlichen
*) Im I. Heft der Beiträge (Alcyonella).
einwärts, in der Ebne des Querschnitts der Tentakeln. Die Erregungswelle schreitet hier an der
rechten Seite empor, an der linken herab. Dadurch müssen die zum Munde verlaufenden Einzelströme
eine leichte Ablenkung nach links erfahren, und der ganze Strudel in eine drehende Bewegung versetzt
werden, die durch die Hufeisenform der Tentakelkrone noch mehr complicirt wird. Das zusammengetriebene
Wasser fliesst an der Basis der Tentakeln, zwischen denselben hindurch, wieder nach aussen,
wobei durch den reusenartig wirkenden Apparat der seitlichen Cilien und durch die Kelchmembran die
Nahrungstheilchen zurückgehalten werden. Die grossen lateralen Cilien lassen sich durch den Leib der
Zelle bis in die Nähe der Stützmembran verfolgen. An der Vorder- und Rückseite der Tentakeln
finden sich bei allen Süsswasserformen jene starren Tastfäden, welche Nitsche zuerst bei Alcyonella
entdeckt hat.
Dass die Tentakelspitze gewisse Besonderheiten zeigt, hat schon Eichhorn*) bemerkt. Nach
ihm befindet sich daselbst „eine Vertiefung“, nach Allman**) „ein kleiner Hohlraum, der von dem
übrigen Theil der Röhre wie abgeschnitten erscheint,“ was nach Nitsche dadurch geschieht, dass die
innére Epithelschicht hier „eine die Höhlung durchsetzende Scheidewand“ (Fig. 102, s) bildet. Ich
habe an lebenden Thieren und an Schnitten wiederholt zu erkennen geglaubt, dass diese obere Kammer
durch eine an ihrer äussersten Spitze gelegene Oeffnung mit der Aussenwelt in Verbindung stehe. Die
Oeffnung erschien viel breiter, wenn der Tentakel seitwärts, als wenn er von vorn oder vom Rücken
betrachtet wurde, so dass sie ein spaltförmiges, der Symmetrieebne des Tentakels parallel gerichtetes
Lumen besitzen muss. Welchem Zweck diese Einrichtung dient, darüber bin ich zu keiner klaren
Vorstellung gelangt. Ein Sinnesorgan zu vermuthen, liegt nahe.
Bei allen PhylaCtolaemen werden die Tentakeln an der Basis durch eine Membran verbunden,
welche zwischen ihnen mehr oder weniger bauschige Falten schlägt und als Kelch oder Intertentakular-
membran bekannt ist (Taf. III, Fig. 46, k); Sie besteht aus einer doppelten Schicht äusserst niedriger
Ectodermzellen, denen eine Fortsetzung der homogenen Membran zur Grundlage dient (Nitsche). Die-
Zellen der Innenseite, welche dem Mundepithel angehören, scheinen von besonders zarter Structur zu
sein, da ich an conservirten Exemplaren von Cristatella fast nur ihre Grenzen erkennen konnte, die sieh
als feine häutige Rippen auf der Membran erhoben, während der Leib grösstentheils geschwunden war.
Die sehr energischen Contractionen der Kelchfalten, welche am lebenden Thier zuweilen beobachtet
werden, lassen auf Querfasern schliessen, die ich bei Fredericella und Plum. fruticosa auch wirklich zu
sehen glaubte.
Die Membran wird dadurch gebildet, dass die ganze Leiste des Lophophorrandes, auf der die
Tentakeln entstehen, schliesslich als Duplicatur mit dieser emporgehoben und zu einer dünnen Lamelle
ausgezogen wird. Bei Cristatella und Plum. fruticosa verschmelzen die benachbarten Lateraltheile der
Kelchfalten mit einander, so dass dann eine continuirliche Aussenlamelle durch Quersepten mit den
Tentakeln verbunden ist (vgl. Taf. V, Fig. 63, XI). Bei Fredericella, Plum. repens und fungosa ist das
nicht der Fall (Fig. 64, V). Ueber Plum. emarginata und vesicularis habe ich nichts notirt.
*) Beiträge zur Naturgeschichte der kleinsten Wasserthiere S. 45.
**) Monograph S. 21.