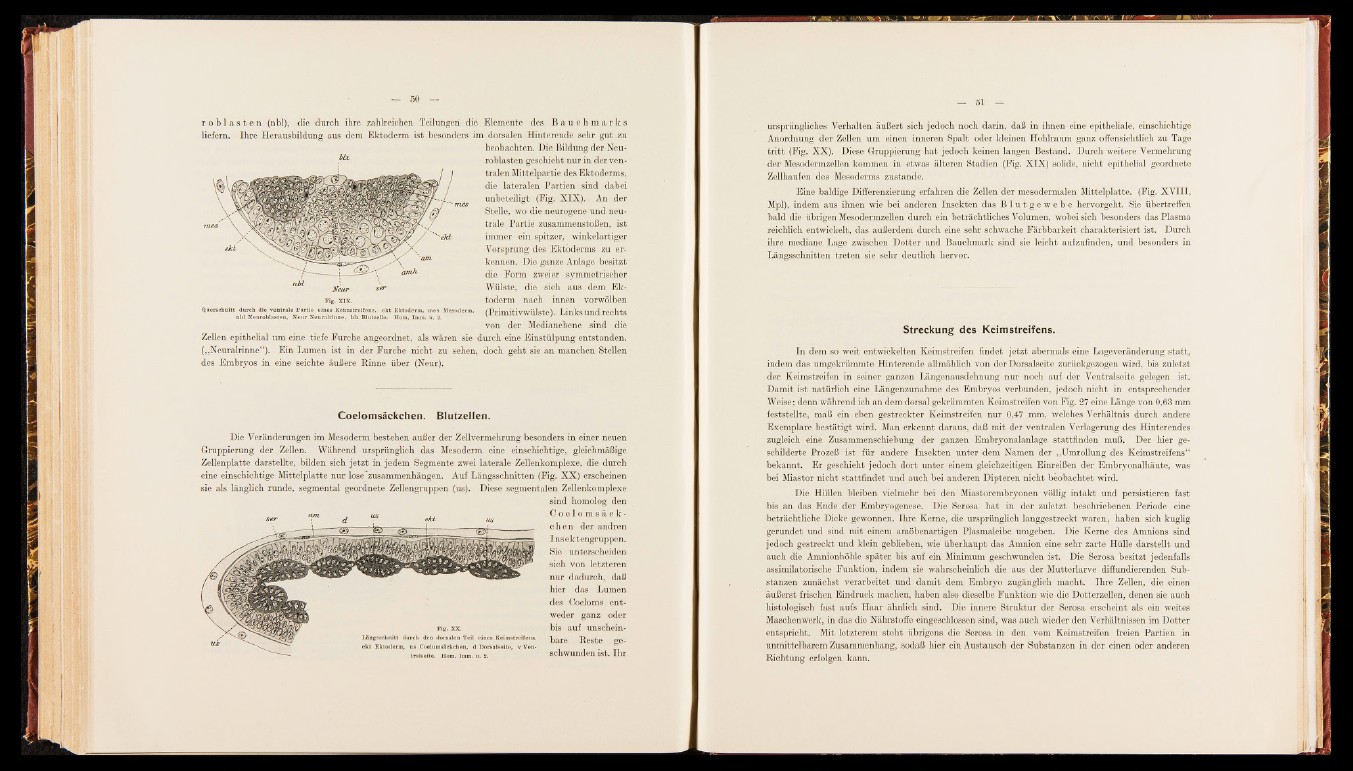
r o b l a s t e n (nbl), die durch, ihre zahlreichen Teilungen die Elemente des B a u c h m a r k s
liefern. Ihre Herausbildung aus dem Ektoderm ist besonders im dorsalen Hinterende sehr gut zu
beobachten. Die Bildung der Neu-
; roblasten geschieht nur in der ventralen
Mittelpartie des Ektoderms,
die lateralen Partien sind dabei
unbeteiligt mes & v( Fi6g . XIX)' . An der
Stelle, wo die neurogene und neutrale
Partie zusammenstoßen, ist
immer ein spitzer, winkelartiger
Vorsprung des Ektoderms zu erkennen.
Die ganze Anlage besitzt
die Form zweier symmetrischer
Wülste, die sich aus dem Ek-
Fig. xj x. toderm nach innen vorwölben
Querschnitt durch die ventrale Partie eines Keimstreifens, ekt Ektoderm, mes Mesoderm, (PrimitivWÜlste) Links Und rechts
nbl Neuroblasten, Neur Neuralrinne, blz Blutzelle. Hom, Imm. u . 2. ' ' ' *
von der Medianebene sind die
Zellen epithelial um eine tiefe Furche angeordnet, als wären sie durch eine Einstülpung entstanden.
(„Neuralrinne“). Ein Lumen ist in der Furche nicht zu sehen, doch, geht sie an manchen Stellen
des Embryos in eine seichte äußere Rinne über (Neur).
Coelomsäckchen. Blutzellen.
Die Veränderungen im Mesoderm bestehen außer der Zellvermehrung besonders in einer neuen
Gruppierung der Zellen. Während ursprünglich das Mesoderm eine einschichtige, gleichmäßige
Zellenplatte darstellte, bilden sich jetzt in jedem Segmente zwei laterale Zellenkomplexe, die durch
eine einschichtige Mittelplatte nur lose Zusammenhängen. Auf Längsschnitten (Fig. XX) erscheinen
sie als länglich runde, segmental geordnete Zellengruppen (us). Diese segmentalen Zellenkomplexe
sind homolog den
C o e l o m s ä c k - us chen der andren
Insektengruppen.
Sie unterscheiden
sich von letzteren
nur dadurch, daß
hier das Lumen
des Coeloms entweder
ganz oder
Fig. xx. bis auf unschein-
LSngsscbnitt durch den dorsalen Teil eines Keimstreifens. b a r e R e s t e £G -
ekt Ektoderm, u s Coelumsäckclien, d Dorsalseite, v Ventralseite.
Hom. Imm. u. 2. schwunden ist. Ihr
ursprüngliches Verhalten äußert sich jedoch noch darin, daß in ihnen eine epitheliale, einschichtige
Anordnung der Zellen um einen inneren Spalt oder kleinen Hohlraum ganz offensichtlich zu Tage
tritt (Fig. XX). Diese Gruppierung hat jedoch keinen langen Bestand. Durch weitere Vermehrung
der Mesodermzellen kommen in etwas älteren Stadien (Fig. XIX) solide, nicht epithelial geordnete
Zellhaufen des Mesoderms zustande.
Eine baldige Differenzierung erfahren die Zellen der mesodermalen Mittelplätte. (Fig. XVIII,
Mpl), indem aus ihnen wie bei anderen Insekten das B l u t g e w e b e hervorgeht. Sie übertreffen
bald die übrigen Mesodermzellen durch ein beträchtliches Volumen, wobei sich besonders das Plasma
reichlich entwickelt, das außerdem durch eine sehr schwache Färbbarkeit charakterisiert ist. Durch
ihre mediane Lage zwischen Dotter und Bauchmark sind sie leicht aufzufinden, und besonders in
Längsschnitten treten sie sehr deutlich hervor.
Streckung des Keimstreifens.
In dem so weit entwickelten Keimstreifen findet jetzt abermals eine Lage Veränderung statt,
indem das umgekrümmte Hinterende allmählich von der Dorsalseite zurückgezogen wird, bis zuletzt
der Keimstreifen in seiner ganzen Längenausdehnung nur noch auf der Ventralseite gelegen ist.
Damit ist natürlich eine Längenzunahme des Embryos verbunden, jedoch nicht in entsprechender
Weise; denn während ich an dem dorsal gekrümmten Keimstreifen von Fig. 27 eine Länge von 0,63 mm
feststellte, maß ein eben gestreckter Keimstreifen nur 0,47 mm, welches Verhältnis durch andere
Exemplare bestätigt wird. Man erkennt daraus, daß mit der ventralen Verlagerung des Hinterendes
zugleich eine Zusammenschiebung der ganzen Embryonalanlage stattfinden muß. Der hier geschilderte
Prozeß ist für andere Insekten unter dem Namen der „Umrollung des Keimstreifens“
bekannt. Er geschieht jedoch dort unter einem gleichzeitigen Einreißen der Embryonalhäute, was
bei Miastor nicht stattfindet und auch bei anderen Dipteren nicht beobachtet wird.
Die Hüllen bleiben vielmehr bei den Miastorembryonen völlig intakt und persistieren fast
bis an das Ende der Embryogenese. Die Serosa hat in der zuletzt beschriebenen Periode eine
beträchtliche Dicke gewonnen. Ihre Kerne, die ursprünglich langgestreckt waren, haben sich kuglig
gerundet und sind mit einem amöbenartigen Plasmaleibe umgeben. Die Kerne des Amnions sind
jedoch gestreckt und klein geblieben, wie überhaupt das Amnion eine sehr zarte Hülle darstellt und
auch die Amnionhöhle später bis auf ein Minimum geschwunden ist. Die Serosa besitzt jedenfalls
assimilatorische Funktion, indem sie wahrscheinlich die aus der Mütterlarve diffundierenden Substanzen
zunächst verarbeitet und damit dem Embryo zugänglich macht. Ihre Zellen, die einen
äußerst frischen Eindruck machen, haben also dieselbe Funktion wie die Dotterzellen, denen sie auch
histologisch fast aufs Haar ähnlich sind. Die innere Struktur der Serosa erscheint als ein weites
Maschenwerk, in das die Nährstoffe eingeschlossen sind, was auch wieder den Verhältnissen im Dotter
entspricht. Mit letzterem steht übrigens die Serosa in den vom Keimstreifen freien Partien in
unmittelbarem Zusammenhang, sodaß hier ein Austausch der Substanzen in der einen oder anderen
Richtung erfolgen kann.