V o r g a n g verbunden: In die aus den Polplatten entstehenden Tochterkerne geht nur die halbe
Chromosomenzahl über. Ist dies wirklich der Fall, so muß bei der folgenden Mitose der Somakerne
ebenfalls wieder die reduzierte Chromosomenzähl nachweisbar sein, und auch alle späteren Abkömmlinge
der 14 Furchungskerne dürfen nur den halben Bestand der Chromosomen enthalten. Diese
Folgerung bestätigt sich durchgängig. Denn sowohl in dem Soma des 15-zelligen, 30-zelligen und
58-zelligen Stadiums, als auch in älteren Blastoderm- und Keimstreifenstadien und in den Geweben
differenzierter Embryonen, in Neuroblasten, Fettkörper-, Muskel- und Drüsenzellen wird in den
karyokinetischen Figuren immer nur die Hälfte der ursprünglichen Chromosomenzahl festgestellt.
Ich habe eine große Zahl von Kernspindeln daraufhin studiert und finde in den meisten 10 oder 11
Chromosomen, in Spindeln , mit seitlicher Ansicht, die für Beantwortung der Zahlenfrage weniger
geeignet sind, wurden mehrfach sogar nur 9 oder 8 Chromosomen gezählt;-.¿Da aber 10 und 11 Chromosomen
gerade bei günstigen Polansichten festgestellt wurden, entscheide ich mich für diese Zahlen.
Mehr als 11 Chromosomen werden nicht vorgefunden. Von diesem Ergebnis aus ist auch ein Rückschluß
auf die ursprüngliche Chromosomenzahl gestattet. Die Auszählungen ergaben dort an und für
sich eine Streuung von 20 bis 24, gemäß der reduzierten Chromosomenzahl aber muß die ursprüngliche
Zahl 20 oder 22 sein; und sie wird nicht größer als 22 sein, wenn man nicht etwa im Keimbläschen
und den ersten Furchungskernen das Vorhandensein accessorischer Chromosomen annehmen will.
Wir stehen hier vor der eigenartigen Erscheinung, daß der Diminutionsprozeß und der
Reduktionsprozeß in derselben Karyokinese vereinigt sind, und es erhebt sich die weitere Frage nach
dem M o d u s d i e s e s V o r g a n g e s .
Der Reduktionsprozeß ist sehr eingehend studiert für eine große Zahl von Objekten bei der
Reifung der männlichen und weiblichen Geschlechtszellen. Dabei ergibt sich, daß in den Kernen
der Spermatocyten und Oocyten die Chromosomen paarweise kopulieren zu bivalenten Einheiten
(Tetraden), Jeder Paarling enthält zwei Chromosomen als Komponenten, und die Zahl der Paarlinge
ist daher die Hälfte der ursprünglichen Chromosomenzahl. Die Paarlinge ordnen sich nun bei der
Reduktionsteilung in der Weise, daß ihre Längsrichtung in die Richtung der Spindelfasern fällt und
ihre Verschmelzungsstellen in die Äquatorialebene zu Hegen kommen. In den Verschmelzungsstellen
erfolgt nun wieder die Trennung, die Hälfte der Komponenten wandert darauf nach dem einen Pol,
die andere Hälfte nach dem anderen Pol, und damit ist ja die Reduktion der Chromosomenzahl
erreicht. Es ist wahrscheinlich, daß der Reduktionsprozeß in unserem Falle in der gleichen Weise
vor sich geht. Die Spindelfiguren, wie deren eine in Figur VIII a dargesteilt ist, lassen kaum eine
andere Deutung zu. Auch hier Hegen die Chromosomen in der Mittelplatte sowohl, wie in den Polplatten
in der Richtung der Spindelfasern. Da ferner die Polplatten je 10 oder 11 Segmente enthalten,
müssen auch in der Mittelplatte nach jeder Seite 10 oder 11 Segmente gerichtet sein. Die letzteren
bilden in der Äquatorialebene paarweise scharfe Winkel, und daran erkennt man, daß hier je zwei
Komponenten zusammenstoßen. Die Mittelplatte muß demnach 11 Doppelchromosomen enthalten,
und diese Zahl habe ich tatsächlich in der Mittelplatte der Spindel in Fig. VIII a festgestellt.
Es wäre nun sehr wünschenswert, eine Äquatorialplatte mit den Doppelchromosomen kurz
vor der Abschnürung der Polplatten anzutreffen. Leider aber habe ich ein solches Stadium nicht
vorgefunden. Doch fand ich in 5 achtzelHgen Embryonen die Furchungskerne in noch früheren
Phasen dieser wichtigen Mitose. Fig. 17 veranschauHcht einen ungefähr medianen Längsschnitt von
10 [a Dicke eines solchen Embryos. Von den acht Kernen wurden fünf getroffen, nämHch der
Urgeschlechtskern am hinteren Pol, ein hinterer Furchungskern im Spiremstadium, ein mittlerer
Kern im Stadium der Diakinese und zwei vordere Kerne im Asterstadium. Von den übrigen drei
Kernen befinden sich noch zwei vorn Hegende ebenfalls im Asterstadium und ein hinterer in der
Phase des Spirems. In den vorderen Kernen findet man die Chromosomen noch nicht zur Äquatorialplatte
angeordnet, sondern noch locker im ganzen Raume der sich bildenden Spindeln verteilt,
weswegen hier für das Auszählen der Chromosomen besonders günstige Verhältnisse vorfiegen. Ich
zählte in den vier Spindeln 20, 20, 21 und 22 Chromosomen, was der Normalzahl entspricht. In
einem anderen gleichartigen Embryo, dessen vordere Kerne sich ebenfaUs im Asterstadium befinden,
habe ich in drei Kernen je 20, in einem 21 Chromosomen festgestellt. Man erkennt daraus, daß von
Haus aus in den Kernen des achtzelHgen Stadiums die Normalzahl der Chromosomen vorhanden ist,
und daß eben eine Kopulation je zweier Chromosomen
erfolgen muß, wenn in die Polplatten nur die
Hälfte der Chromosomen (und zwar nur deren
äußere Segmente) übergehen soUen.
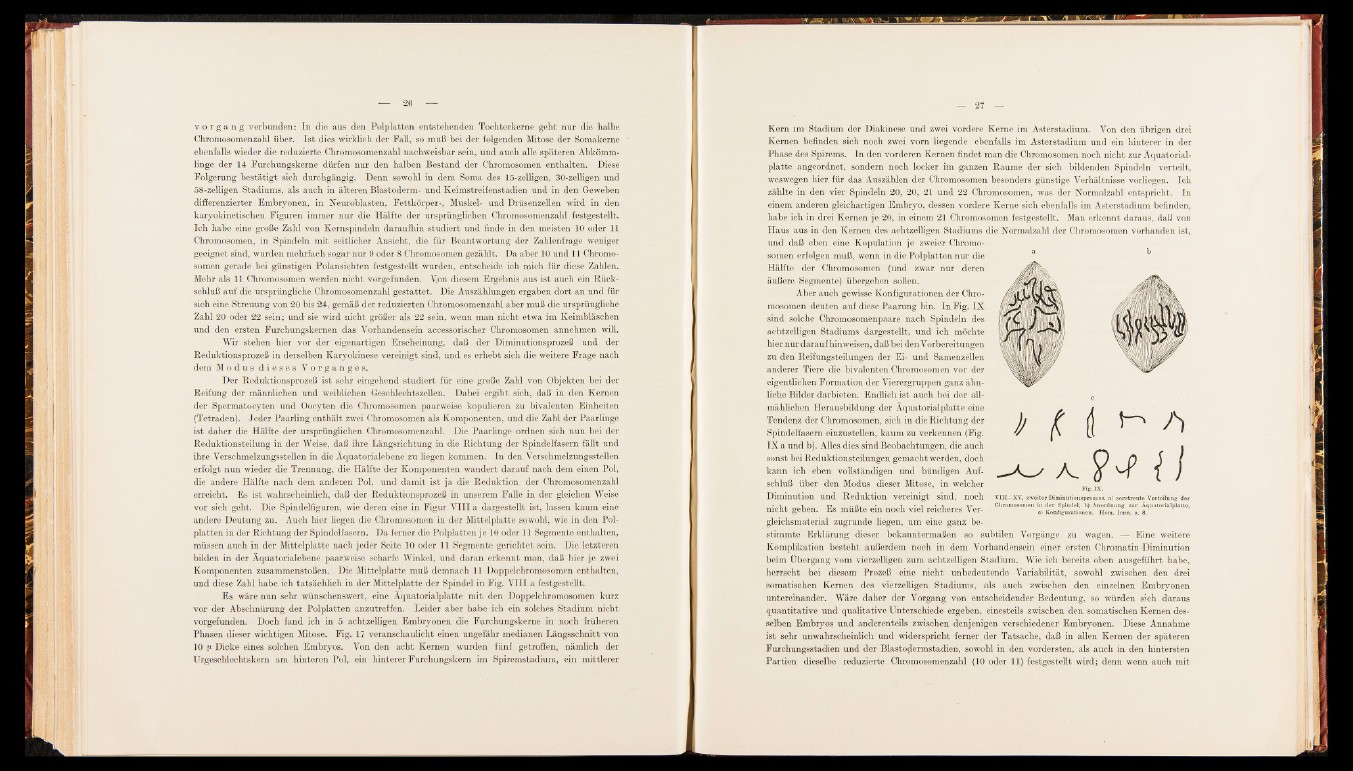
Aber auch gewisse Konfigurationen der Chromosomen
deuten auf diese Paarung hin. In Fig. IX
sind solche Chromosomenpaare nach Spindeln des
achtzelHgen Stadiums dargestellt, und ich möchte
hier nur darauf hin weisen, daß bei den Vorbereitungen
zu den Reifungsteilungen der Ei- und Samenzellen
anderer Tiere die bivalenten Chromosomen vor der
eigentHchen Formation der Vierergruppen ganz ähn-
Hche Bilder darbieten. EndHch ist auch bei der allmählichen
Herausbildung der Äquatorialplatte eine
Tendenz der Chromosomen, sich in die Richtung der
Spindelfasern einzusteUen, kaum zu verkennen (Fig.
IX a und b). Alles dies sind Beobachtungen, die auch
sonst bei Reduktionsteilungen gemacht werden, doch
kann ich eben vollständigen und bündigen Aufschluß
über den Modus dieser Mitose, in welcher
Diminution und Reduktion vereinigt sind, noch
nicht geben. Es müßte ein noch viel reicheres Vergleichsmaterial
zugrunde Hegen, um eine ganz bestimmte
h ( 1 f - A
Fig. IX.
VIII—XV, zweiter Diminutionsprozess, a) zerstreute Verteilung der
Chromosomen in der Spindel, b) Anordnung zur Äquatorialplatte,
' c) Konfigurationen. Hom. Imm. u. 8.
Erklärung dieser bekanntermaßen so subtilen Vorgänge zu wagen. — Eine weitere
KompHkation besteht außerdem noch in dem Vorhandensein einer ersten Chromatin-Diminution
beim Übergang vom vierzelHgen zum achtzelHgen Stadium. Wie ich bereits oben ausgeführt habe,
herrscht bei diesem Prozeß eine nicht unbedeutende VariabiHtät, sowohl zwischen den drei
somatischen Kernen des vierzelHgen Stadiums, als auch zwischen den einzelnen Embryonen
untereinander. Wäre daher der Vorgang von entscheidender Bedeutung, so würden sich daraus
quantitative und quaHtative Unterschiede ergeben, einesteils zwischen den somatischen Kernen desselben
Embryos und anderenteils zwischen denjenigen verschiedener Embryonen. Diese Annahme
ist sehr unwahrscheinHch und widerspricht ferner der Tatsache, daß in aHen Kernen der späteren
Furchungsstadien und der Blastodermstadien, sowohl in den vordersten, als auch in den hintersten
Partien dieselbe reduzierte Chromosomenzahl (10 oder 11) festgesteUt wird; denn wenn auch mit