Nach den in Fig. 10—12 abgebildeten Stadien, die Will freilich als abnorm bezeichnet, könnte man
in der Tat auf eine Einstülpung der Geschlechtszellen mit den davorliegenden Somazellen schließen,
und wäre dies der Fall, so wäre allerdings hier eine fast vollkommene Analogie mit den entsprechenden
Vorgängen bei Miastor vorhanden.
Eine Art Invagination der Geschlechtszellen beschreibt auch Heymons; denn bei Periplaneta
Phyllodromia und Gryllus findet er am Hinterende des Keimstreifens eine Vertiefung, die „Geschlechtsgrube“,
m deren Wand die Geschlechtszellen hegen, und die dadurch entsteht, daß letztere aus der
oberflächlichen Schicht nach innen wandern.
Endlich hat Voeltzkow bei Musca vomitoria gefunden, daß die Polzellen durch ihren Druck
die davorhegenden Blastodermzehen nach innen drängen, daß letztere dann als ein Zapfen in das
Ei hineinragen, sich aber später von einander lösen und Dotterzellen hefem. Dies entspricht genau
den Verhältnissen, die in meinen Figuren 23, 24, 25 veranschaulicht sind.
Zur Beurteilung des hier erörterten Problems halte ich mich zu einer weiteren Mutmaßung
für berechtigt. Sie. bezieht sich auf die chromatischen Bestkörper. Es erscheint mir kaum wahrscheinlich,
daß der Diminutionsprozeß nur bei Miastor stattfinden sollte, vielmehr wird er auch bei
anderen und viellleicht hei allen Insekten vorhanden sein. Dann müßten aber auch im Dotter junger
-Embryonen die Bestkörper wahrgenommen werden. Daß aber letztere bei ihrer tiefen Färbung
leicht für Kerne genommen werden können, liegt auf der Hand, und ich habe selbst diesen Irrtum
begangen, bevor ich den Diminutionsprozeß kannte. Man vergleiche hierzu, was Gräber (22) über
die Dotterzellen in seinen „Vergleichenden Studien über die Embryologie der Insekten“ :sagt: „Ferner
sind die Vitelloblasten der Musciden meiner Ansicht nach wirklich keine eigentlichen Zellen zu nennen,
sondern im allgemeinen bloße, amöboid zerfließende Kerne.“ In Hunderten derselben, die er- mit
den besten Linsen durchmusterte, fand er nie „etwas einem Kern Vergleichbares“. In toto erscheinen
sie stets weit intensiver tingiert, als die Leiber der Gewebszellen und enthalten im Inneren zahlreiche,
größere und kleinere Chromatinkörperchen. Dazu kommt noch, „daß um die zum Teil den Proto-
blästencharakter beibehaltenden Biesenkerne hier meistens gar kein Protoplasmarest nachweisbar
ist, eine Erscheinung, die aber bekanntlich auch bei anderen Insekten z.B. teilweise vonGrassi (25)
hei der Biene beobachtet wurde“. Endlich hat Gräber auch keine Übergänge von den Vitelloblasten
zu den echten Gewebszellen finden können. Das sind alles Angaben, die eigentlich-gar nicht auf
Dotterzellen, sondern nur auf chromatische Bestkörper schließen lassen. Man betrachte hierzu die
Figuren 6, Tf. I und 69, Tf. VI der oben genannten Abhandlung und wird darin die fast frappante
Ähnlichkeit der angeblichen Dotterkerne mit den Chromatinresten von Miastor erkennen. Ähnliche
Mutmaßungen lassen sich auch an Fig. 8 der Will’schen Aphidenarbeit anknüpfen. Die kleiner, Körper
im Inneren gleichen gar nicht den sonst abgebildeten Dotterzellen. Außerdem ist Wills Ansicht
höchst unwahrscheinlich, daß die Bildung des Entoderms zu ganz verschiedener Zeit beginnen könne.
Dieser Prozeß wird jedenfalls auch hier an ein ganz bestimmtes Stadium (vielleicht an das der Fig. 9)
gebunden sein, und die inneren Zellen der Eguren 4—7 würden dann zurückgebliebenen Furchungszellen
entsprechen, während ich in dem angeblichen Entoderm der Fig. 8 Chromatinreste vermute.
Auch noch in andren Insektenarbeiten ist es auffällig, daß die Dotterzellen häufig als äußerst hinfällige
Gebilde beschrieben werden. Ihre Bildung widerspräche dann dem ökonomischen Prinzip. Bei
Miastor fand ich im Gegenteil, daß sie während eines großen Teils der Embryonalentwicklung einen
äußerst aktiven Eindruck erwecken. Erst mit dem Beginn der Gewebsdifferenzierung fangen einzelne
an, zu degenerieren.
Und sollten vielleicht hiermit auch die rätselhaften Parac^ten Heymons’ ihre Erklärung
finden? Heymons (36) hält sie von.den Dotterzellen scharf auseinander und beschreibt an ihnen
ganz charakteristische Degenerationserscheinungen. Man vergleiche hierzu besonders die Figur 42!
Daß sie aber mit Chromatinresten identisch seien, soll hier ebenfalls nur als Vermutung ausgesprochen
werden.
Wachstum des Keimstreifens. Embryonalhüllen.
Nach diesen Betrachtungen wenden wir uns wieder zu der weiteren Entwicklung unserer
Miastor-Embryonen. An dem einmal angelegten Keimstreifen gehen in rascher Folge weitere
Veränderungen vor sich. Vor allen Dingen findet jetztt ein starkes Wachstum statt. Der Embryo
wird größer und größer, der Keimstreifen findet aber trotzdem auf der Ventralseite keinen Platz
mehr und krümmt sich deshalb .am hinteren Pol nach der Dörsalseite um, während der Kopfteil des
Embryos seine Lage unverändert beibehält. Das umgekrümmte Hinterende gewinnt immer mehr
an Ausdehnung und gelangt dadurch längs des Kückens immer weiter nach vom, bis es zuletzt nur
noch um ein kleines Stück von den, Scheitellappen entfernt ist (Eg. 27)., Wie bekannt, lagen die Keimbahnzellen
nach ihrer Invagination dem Hinterende des Keimstreifens auf (Fig. 24, 25). Diese Lage
ändern sic auch bei dem nun folgenden Wachstum des, Keimstreifens nicht, sondern sie werden dabei
Tor lhm hergeschoben und sind deshalb in jeder Phase am äußersten Ende des dorsalen Keimstreifenabschnitts
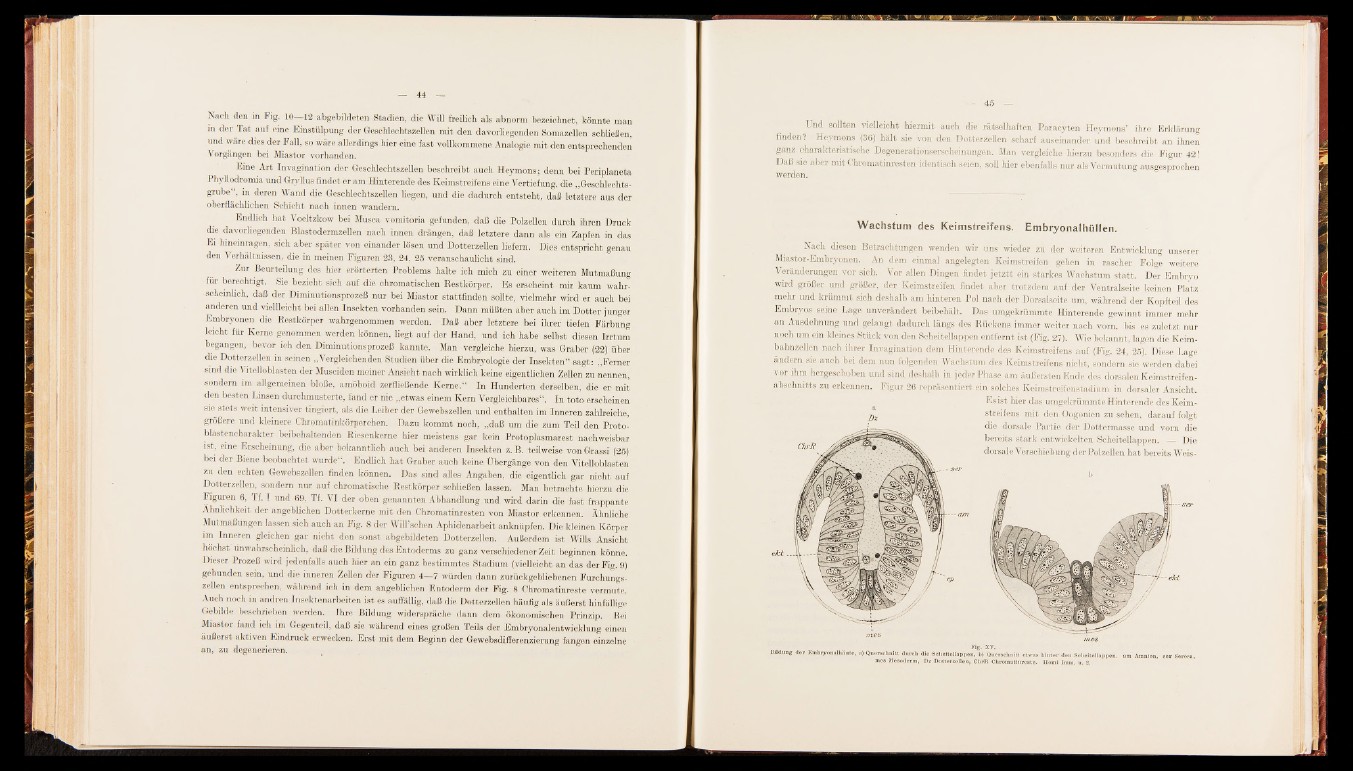
zu erkennen. Figur 26 repräsentiert ein solches Keimstreifenstadium in dorsaler Ansicht
Es ist hier das umgekrümmte Hinterende des Keim-
j)x Streifens mit den Oogonien zu sehen, darauf folgt
die dorsale Partie der Dottermasse und vorn die
bereits stark entwickelten Scheitellappen. — Die
dorsale Verschiebung der Polzellen hat bereits Weis
fflfts
Fig. XV. .
Bildung d e r Em b ry o n alh äu te , a) Q u e rsc h n itt d u rch die Soh e ite llap p en , b) Q u e rsc h n itt e twas h in te r d e n Sch e itellap p en , am 'Am n io n , s e r Se rosa,
mes Mesoderm, Dz D o tte rz ellen , ChrR C h rom a tin reste. Homi Imm. u . 2.