dagegen in ihrem Handbuch die Pneumatophore resp. die primäre Schwimmglocke für das primäre
Individuum, den Polyp bezw. Saugmagen für das distale Ende des aboralen Stolo prolifer, also für
ein Produkt des ersteren. W o l t e r e c k (1905, p. 612) leugnet dies glatt und sieht, ähnlich wie
L e u c k a r t, umgekehrt im Primärpolyp das erste Produkt der Planula, d. h. nach ihm wird die
Planula selbst zum Primärpolypen,
der am aboralen Pol
eine stoloartige Proliferationszone
entwickelt, die dann die
Pneumatophore resp. das lar-
vale Deckstück und später die
weiteren larvalen Deekstücke
und die Schwimmglocken hervorbringt:
„Die medusoide
Luftflasche,“ schreibt er, „ist
das erste, vom Primärpolyp
aboral geknospte Sekundär-
zoid. Also kann unmöglich
umgekehrt der Primärpolyp
als aboral produziertes Stolo-
Ende seines eigenen Geschöpfes,
der Terminalmeduse,
aufgefaßt werden.“
Ugii
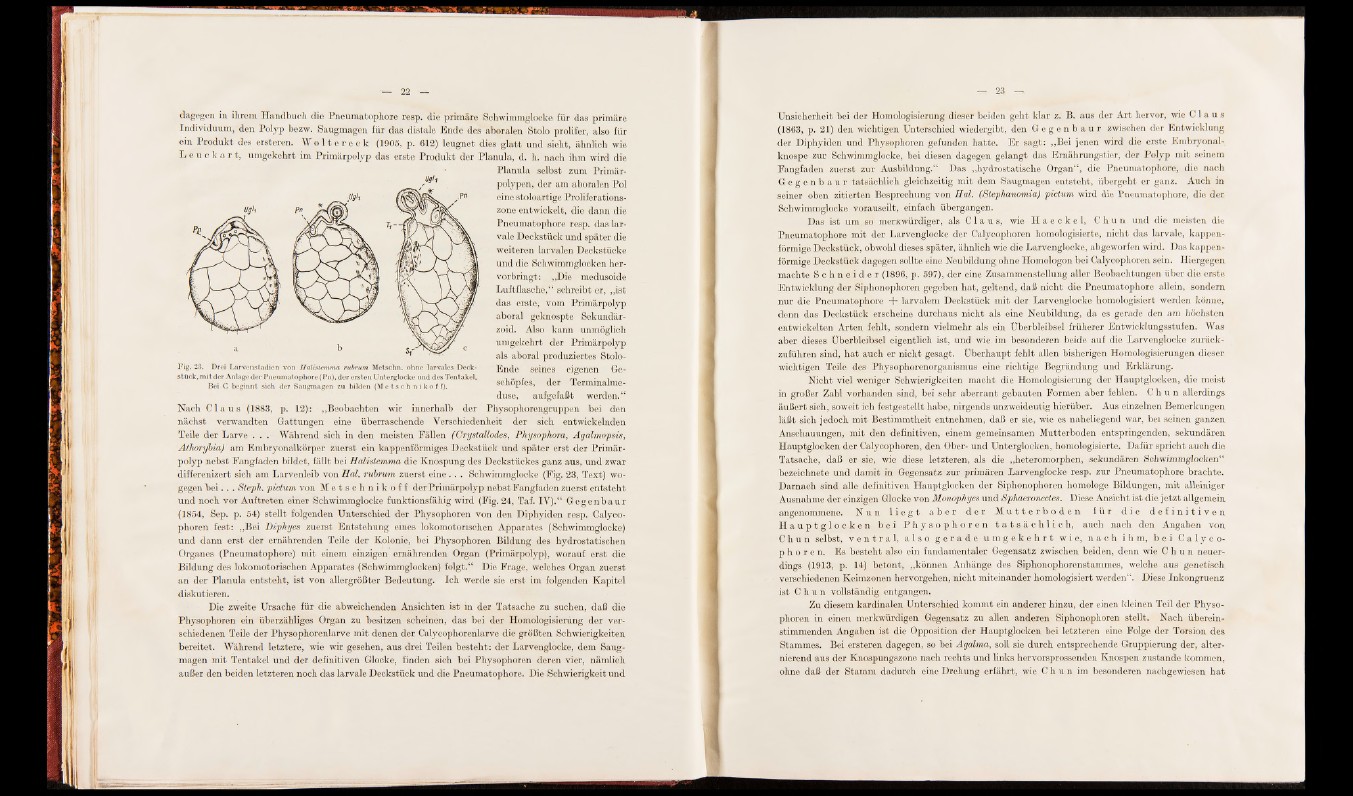
Fig. 23. Drei Larvenstadien von Halistemma rubrum Metschn. ohne larvales Deckstück,
mit der Anlage der Pneumatophore (Pn), der ersten Unterglocke und des Tentakel.
Bei C beginnt sich der Saugmagen zu bilden (Me t s c h n i k o f f ) .
Nach C l a u s (1883, p. 12): „Beobachten wir innerhalb der Physophorengruppen bei den
nächst verwandten Gattungen eine überraschende Verschiedenheit der sich entwickelnden
Teile der Larve . . . Während sich in den meisten Fällen (Crystattodes, Physophora, Agalmopsis,
Athorybia) am Embryonalkörper zuerst ein kappenförmiges Deckstück und später erst der Primärpolyp
nebst Fangfaden bildet, fällt bei Halislemma die Knospung des Deckstückes ganz aus, und zwar
differenizert sich am Larvenleib von Hol. rubrum zuerst eine . . . Schwimmglocke (Fig. 23, Text) wogegen
bei . . . Steph. pictum von M e t s c h n i k o f f der Primärpolyp nebst Fangfaden zuerst entsteht
und noch vor Auftreten einer Schwimmglocke funktionsfähig wird (Fig. 24, Taf. IV).“ Gegenba ur
(1854, Sep. p. 54) stellt folgenden Unterschied der Physophoren von den Diphyiden resp. Calyco-
phoren fest: „Bei Diphyes zuerst Entstehung eines lokomotorischen Apparates (Schwimmglocke)
und dann erst der ernährenden Teile der Kolonie, bei Physophoren Bildung des hydrostatischen
Organes (Pneumatophore) mit einem einzigen ernährenden Organ (Primärpolyp), worauf erst die
Bildung des lokomotorischen Apparates (Schwimmglocken) folgt.“ Die Frage, welches Organ zuerst
an der Planula entsteht, ist von allergrößter Bedeutung. Ich werde sie erst im folgenden Kapitel
diskutieren.
Die zweite Ursache für die abweichenden Ansichten ist in der Tatsache zu suchen, daß die
Physophoren ein überzähliges Organ zu besitzen scheinen, das bei der Homologisierung der verschiedenen
Teile der Physophorenlarve mit denen der Calycophorenlarve die größten Schwierigkeiten
bereitet. Während letztere, wie wir gesehen, aus drei Teilen besteht: der Larvenglocke, dem Saugmagen
mit Tentakel und der definitiven Glocke, finden sich bei Physophoren deren vier, nämlich
außer den beiden letzteren noch das larvale Deckstück und die Pneumatophore. Die Schwierigkeit und
Unsicherheit bei der Homologisierung dieser beiden geht klar z. B. aus der Art hervor, wie C l a u s
(1863, p. 21) den wichtigen Unterschied wiedergibt, den G e g e n b a u r zwischen der Entwicklung
der Diphyiden und Physophoren gefunden hatte. Er sagt: „Bei jenen wird die erste Embryonalknospe
zur Schwimmglocke, bei diesen dagegen gelangt das Ernährungstier, der Polyp mit seinem
Fangfaden zuerst zur Ausbildung.“ Das „hydrostatische Organ“, die Pneumatophore, die nach
G e g e n b a u r tatsächlich gleichzeitig mit dem Saugmagen entsteht, übergeht er ganz. Auch in
seiner oben zitierten Besprechung von Hai. (Stephanomia) pictum wird die Pneumatophore, die der
Schwimmglocke vorauseilt, einfach übergangen.
Das ist um so merkwürdiger, als Cl a u s , wie H a e c k e l , C h u n und die meisten die
Pneumatophore mit der Larvenglocke der Calycophoren homologisierte, nicht das larvale, kappenförmige
Deckstück, obwohl dieses später, ähnlich wie die Larvenglocke, abgeworfen wird. Das kappenförmige
Deckstück dagegen sollte eine Neubildung ohne Homologon bei Calycophoren sein. Hiergegen
machte S c h n e i d e r (1896, p. 597), der eine Zusammenstellung aller Beobachtungen über die erste
Entwicklung der Siphonophoren gegeben hat, geltend, daß nicht die Pneumatophore allein, sondern
nur die Pneumatophore + larvalem Deckstück mit der Larvenglocke homologisiert werden könne,
denn das Deckstück erscheine durchaus nicht als eine Neubildung, da es gerade den am höchsten
entwickelten Arten fehlt, sondern vielmehr als ein Überbleibsel früherer Entwicklungsstufen. Was
aber dieses Überbleibsel eigentlich ist, und wie im besonderen beide auf die Larvenglocke zurückzuführen
sind, hat auch er nicht gesagt. Überhaupt fehlt allen bisherigen Homologisierungen dieser
wichtigen Teile des Physophorenorganismus eine richtige Begründung und Erklärung.
Nicht viel weniger Schwierigkeiten macht die Homologisierung der Hauptglocken, die meist
in großer Zahl vorhanden sind, bei sehr aberrant gebauten Formen aber fehlen. C h u n allerdings
äußert sich, soweit ich festgestellt habe, nirgends unzweideutig hierüber. Aus einzelnen Bemerkungen
läßt sich jedoch mit Bestimmtheit entnehmen, daß er sie, wie es naheliegend war, bei seinen ganzen
Anschauungen, mit den definitiven, einem gemeinsamen Mutterboden entspringenden, sekundären
Hauptglocken der Calycophoren, den Ober- und Unterglocken, homologisierte. Dafür spricht auch die
Tatsache, daß er sie, wie diese letzteren, als die „heteromorphen, sekundären Schwimmglocken“
bezeichnete und damit in Gegensatz zur primären Larvenglocke resp. zur Pneumatophore brachte.
Darnach sind alle definitiven Hauptglocken der Siphonophoren homologe Bildungen, mit alleiniger
Ausnahme der einzigen Glocke von Monophyes und Sphaeronectes. Diese Ansicht ist die jetzt allgemein
angenommene. N u n l i e g t a b e r d e r M u t t e r b o d e n f ü r d i e d e f i n i t i v e n
H a u p t g l o c k e n b e i P h y s o p h o r e n t a t s ä c h l i c h , auch nach den Angaben von.
C h u n selbst, v e n t r a l , a l s o g e r a d e , u m g e k e h r t wie, n a c h i hm, b e i C a l y c o p
h o r e n . Es besteht also ein fundamentaler Gegensatz zwischen beiden, denn wie C h u n neuerdings
(1913, p. 14) betont, „können Anhänge des Siphonophorenstammes, welche aus genetisch
verschiedenen Keimzonen hervorgehen, nicht miteinander homologisiert werden“. Diese Inkongruenz
ist C h u n vollständig entgangen.
Zu diesem kardinalen Unterschied kommt ein anderer hinzu, der einen kleinen Teil der Physophoren
in einen merkwürdigen Gegensatz zu allen anderen Siphonophoren stellt. Nach übereinstimmenden
Angaben ist die Opposition der Hauptglocken bei letzteren eine Folge der Torsion des
Stammes. Bei ersteren dagegen, so bei Agalma, soll sie durch entsprechende Gruppierung der, alternierend
aus der Knospungszone nach rechts und links hervorsprossenden Knospen zustande kommen,
ohne daß der Stamm dadurch eine Drehung erfährt, wie C h u n im besonderen nachgewiesen hat