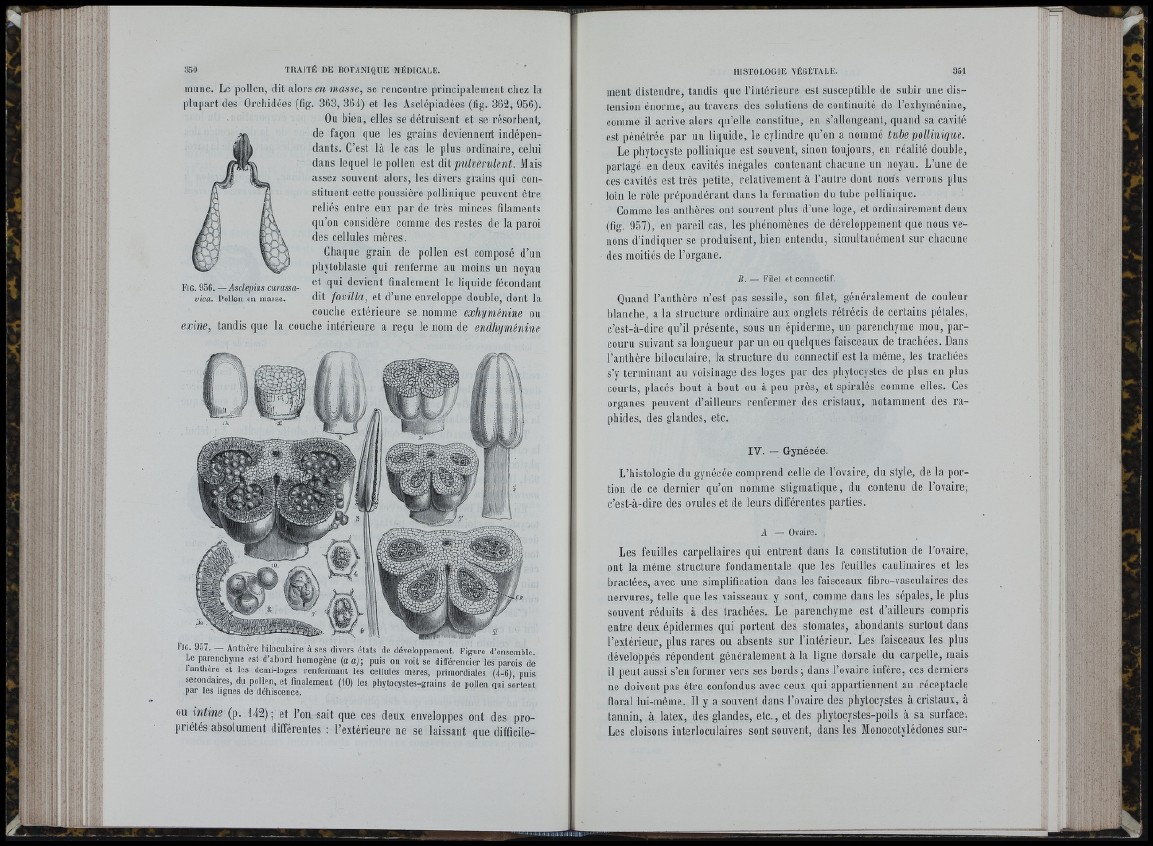
mime. Le pollen, dit alors en masse, se rencontre principalement chez la
plupart des Orchidées (fig. 363, 364) et les Asclépiadées (fig. 362, 956).
Ou bien, elles se détruisent et se résorbent,
de façon que les grains deviennent indépendants.
C’est là le cas le plus ordinaire, celui
dans lequel le pollen est dit pulvérulent. Mais
assez souvent alors, les divers grains qui constituent
celte poussière pollinique peuvent être
reliés entre eux par de très minces filaments
qu’on considère comme des restes de la paroi
des cellules mères.
Chaque grain de pollen est composé d’un
phytoblaste qui renferme au moins un noyau
et qui devient finalement le liquide fécondant
dit fovilla, et d’une enveloppe double, dont la
couche extérieure se nomme exhyménine ou
Fig. 956. — Asclepias curassa-
vica. Pollen en masse.
exine, tandis que la couche intérieure a reçu le nom de endhyménine
l‘ JG. 907. — Anthère biloculaire à ses divers états de développement. Figure d’ensemble.
Le parenchyme est d’abord homogène {a a)-, puis on voit se différencier les parois de
1 anthère et les demi-loges renfermant les cellules mères, primordiales (4-6), puis
secondaires, du pollen, et finalement (10) les phytocystes-grains de pollen qui sortent
par les lignes de déhiscence.
OU intine (p. 142); et Ton sait que ces deux enveloppes ont des propriétés
absolument différentes : l’extérieure ne se laissant que difficilement
distendre, tandis que l’intérieure est susceptible de subir une distension
enorme, au travers des solutions de continuité de l’exhyménine,
comme il arrive alors qu’elle constitue, en s’allongeant, quand sa cavité
est pénétrée par un liquide, le cylindre qu’on a nommé tube pollinique.
Le phytocyste pollinique est souvent, sinon toujours, en réalité double,
partagé en deux cavités inégales contenant cbacune un noyau. L’une de
ces cavités est très petite, relativement à l ’autre dont nous verrons plus
loin le rôle prépondérant dans la formation du tube pollinique.
Comme les anthères ont souvent plus d’une loge, et ordinairement deux
(fig. 957), en pareil cas, les pliénomènes de développement que nous venons
d’indiquer se produisent, bien entendu, simultanément sur cbacune
des moitiés de l’organe.
B. — Filet et connectif.
Quand l’anthère n’est pas sessile, son filet, généralement de couleur
blanche, a la structure ordinaire aux onglets rétrécis de certains pétales,
c’est-à-dire qu’il présente, sous un épiderme, un parenchyme mou, parcouru
suivant sa longueur par un ou quelques faisceaux de trachées. Dans
l’anthère biloculaire, la structure du connectif est la même, les trachées
s’y terminant au voisinage des loges par des phytocystes de plus en plus
courts, placés bout à bout ou à peu près, et spiralés comme elles. Ces
organes peuvent d’ailleurs renfermer des cristaux, notamment des ra phides,
des glandes, etc.
IV. — Gynécée.
L’histologie du gynécée comprend celle de l’ovaire, du style, de la portion
de ce dernier qu’on nomme stigmatique, du contenu de l’ovaire,
c’est-à-dire des ovules et de leurs différentes parties.
A — Ovaire.
Les feuilles carpellaires qui entrent dans la constitution de l’ovaire,
ont la même structure fondamentale que les feuilles caulinaires et les
bractées, avec une simplification dans les faisceaux fibro-vasculaires des
nervures, telle que les vaisseaux y sont, comme dans les sépales, le plus
souvent réduits à des trachées. Le parenchyme est d’ailleurs compris
entre deux épidermes qui portent des stomates, abondants surtout dan^
l ’extérieur, plus rares ou absents sur l’intérieur. Les faisceaux les plus
développés répondent généralement à la ligne dorsale du carpelle, mais
il peut aussi s’en former vers ses bords; dans l’ovaire infère, ces derniers
ne doivent pas être confondus avec ceux qui appartiennent au réceptacle
floral lui-même. Il y a souvent dans l’ovaire des pbytocystes à cristaux, à
tannin, à latex, des glandes, etc., et des phytocystes-poils à sa surface.
Les cloisons interloculaires sont souvent, dans les Monocotylédones sur