und linkem Cardiocoel verschwindet, treten beide „Halshöhlen“ in
offene Kommunikation (Fig. 472) und bilden an der Bauchseite
des Amniotenkopfes einen hufeisenförmigen Bogen, dessen beide
Schenkel sich dorsalwärts nach hinten in die Pleurodukte oder
Pleuralhöhlen fortsetzen und von da aus weiter in die paarigen
Peritonealtaschen des Rumpfes. Aber auch nachdem die beiden
Halshöhlen bereits zur Vereinigung gelangt sind (Fig. 472), liegen
anfänglich die paarigen Herzschläuche noch getrennt, und selbst
nachdem I sie verschmolzen sind,
deutet noch eine zarte Scheidewand
in der Mitte des inneren
einfachen Endothelschlauches
(Fig. 469 s, 473 h) die ursprünglich
getrennte Anlage an. Dieses
cenogenetische „primäre Cardial-
septum“ verschwindet bald und
hat gar keine Beziehung zu der
späteren bleibenden Scheidewand
beider Herzhälften, welche als
Erbstück von den Reptilien eine
hohe palingenetische Bedeutung
beanspruchte
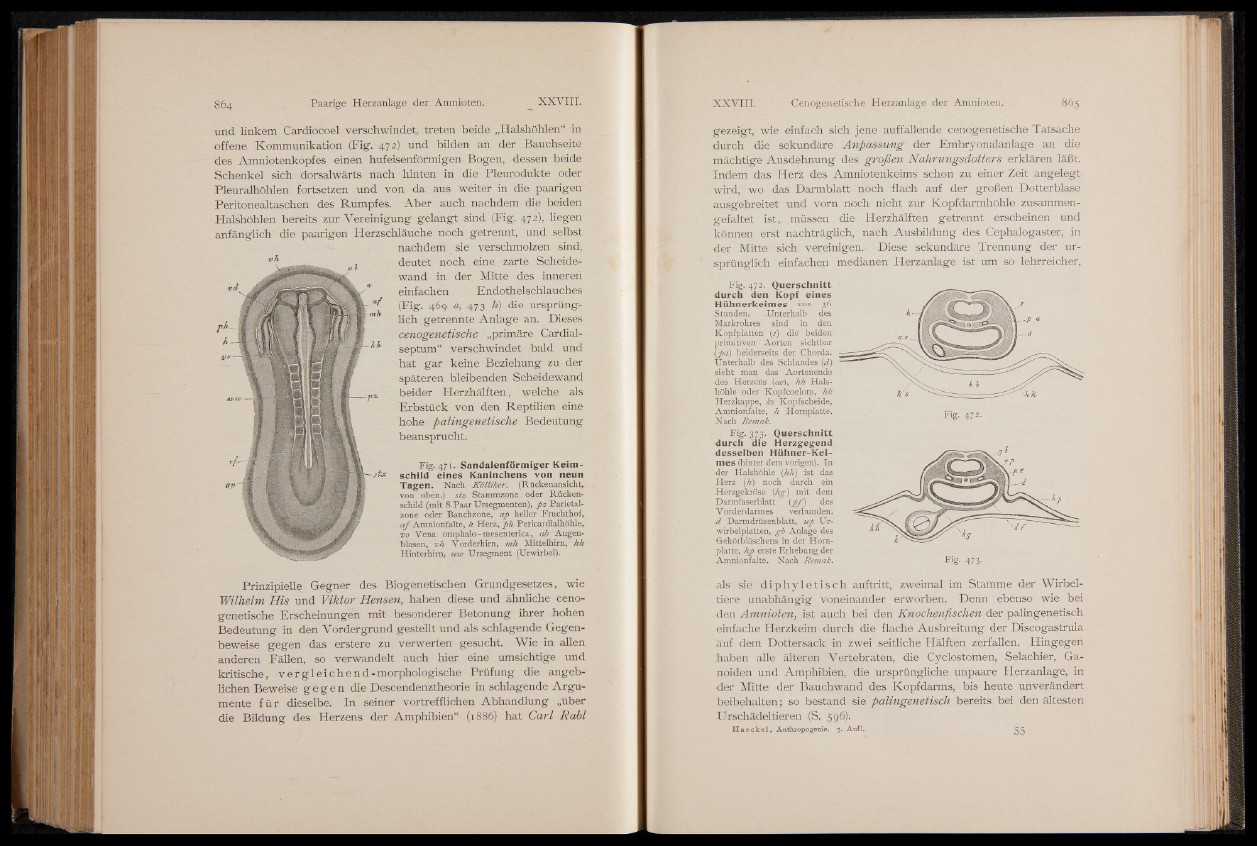
Fig. 471. Sandalenförmiger Keim-
schild eines Kaninchens von neun
Tagen. Nach K ö llik er. (Rückenansicht,,
von oben.) stz Stammzone oder Rückenschild
(mit 8 Paar TJrsegmenten), fiz Parietalzone
oder Bauchzone, afi heller Fruchthof,
a f Amnionfalte, h Herz, fih Pericardialhöhle,
vo Vena omphalo-mesenterica, ab "Augenblasen,
vh Vorderhim, mh Mittelhim, hh
Hinterhim, uw Ursegment (Urwirbel).
Prinzipielle Gegner des Biogenetischen Grundgesetzes, wie
Wilhelm His und Viktor Hensen, haben diese und ähnliche cenogenetische
Erscheinungen mit besonderer Betonung ihrer hohen
Bedeutung in den Vordergrund gestellt und als schlagende Gegenbeweise
gegen das erstere zu verwerten gesucht. Wie in allen
anderen Fällen, so verwandelt auch hier eine umsichtige und
kritische, vergleichend-morphologische Prüfung die angeblichen
Beweise g e g en die Descendenztheorie in schlagende Argumente
fü r dieselbe. In seiner vortrefflichen Abhandlung „über
die Bildung des Herzens der Amphibien“ (1886) hat Carl Rabl
gezeigt, wie einfach sich jene auffallende cenogenetische Tatsache
durch die sekundäre Anpassung der Embryonalanlage an die
mächtige Ausdehnung des großen Nahrungsdotters erklären läßt.
Indem das Herz des Amniotenkeims schon zu einer Zeit angelegt
wird, wo das Darmblatt noch flach auf der großen Dotterblase
.ausgebreitet und vorn noch nicht zur Kopfdarmhöhle zusammengefaltet
ist, müssen die Herzhälften getrennt erscheinen und
können erst nachträglich, nach Ausbildung des Cephalogaster, in
der Mitte sich vereinigen. Diese sekundäre Trennung der ursprünglich
einfachen medianen Herzanlage ist um so lehrreicher,
Fig. 472. Querschnitt
durch den Kopf eines
Hühnerkeimes von 36
Stunden. -Unterhalb des
Markrohres sind in den
Kopfplatten (s) die beiden
primitive^' Aorten sichtbar
Ip a ) beiderseits der Chorda.
Unterhalb des Schlundes (d)
sieht man das Aortenende
■ des Herzens (ae), hh Hals-
höhle oder Kopfcoelom, hk
Herzkappe, ks Kopfscheide,
Amnionfalte, h Hornplatte.
"Hach Remak.
Fig. 472.
Fig-,373* Querschnitt
durch die Herzgegend
desselben Hühner-Keimes
(hinter dem vorigen). In
•der Halshöhle (hJi) ist das
Herz (h)- noch durch ein
Herzgekröse {hg) mit dem
Darmfaserblatt ipf ) des
V orderdarmes verbunden.
d Darmdrüsenblatt, ufi Ur-
wirbelplatten, gb Anlage des
Gehörbläschens in der Horn-
platte, hp erste Erhebung der
Amnionfalte. Nach Remak. Fig- 473-
.als sie diphy le t i s ch auftritt, zweimal im Stamme der Wirbeltiere
unabhängig voneinander erworben. Denn ebenso wie bei
■ den Amnioten, ist auch bei den Knochenfischen der palingenetisch
•einfache Herzkeim durch die flache Ausbreitung der Discogastrula
auf dem Dottersack in zwei seitliche Hälften zerfallen. Hingegen
haben alle älteren Vertebraten, die Cyclostomen, Selachier, Ga-
noiden und Amphibien, die ursprüngliche unpaare Herzanlage, in
der Mitte der Bauchwand des Kopfdarms, bis heute unverändert
beibehalten; so bestand sie palingenetisch bereits bei den ältesten
Urschädeltieren (S. 596).' :
H a e c k e l, Anthropogenie. 5. Aufl.