unterscheiden sind. Beim a-Organ trifft das nur für die Cf cf zu, während bei den 22 durch
die Bildung von Infektionsmaterial in den anschwellenden Infektionshügeln eine weitere
Größenzunahme eintritt, wie überhaupt bei den a-Organen schon vom III. Larvenstadium
(Fig. 26) an ein Geschlechtsdimorphismus bezüglich der Größe zu bemerken ist, der bei den
X-Organen völlig fehlt. Dieser Unterschied ist leicht verständlich; denn das Material (In-
fektionssymbionten),das bei den weiblichen a-Organen ein gesteigertes Wachstum gegenüber
den männlichen Mycetomen bedingt, ist ja beim X-Organ schon auf embryonalen Stadien
a e
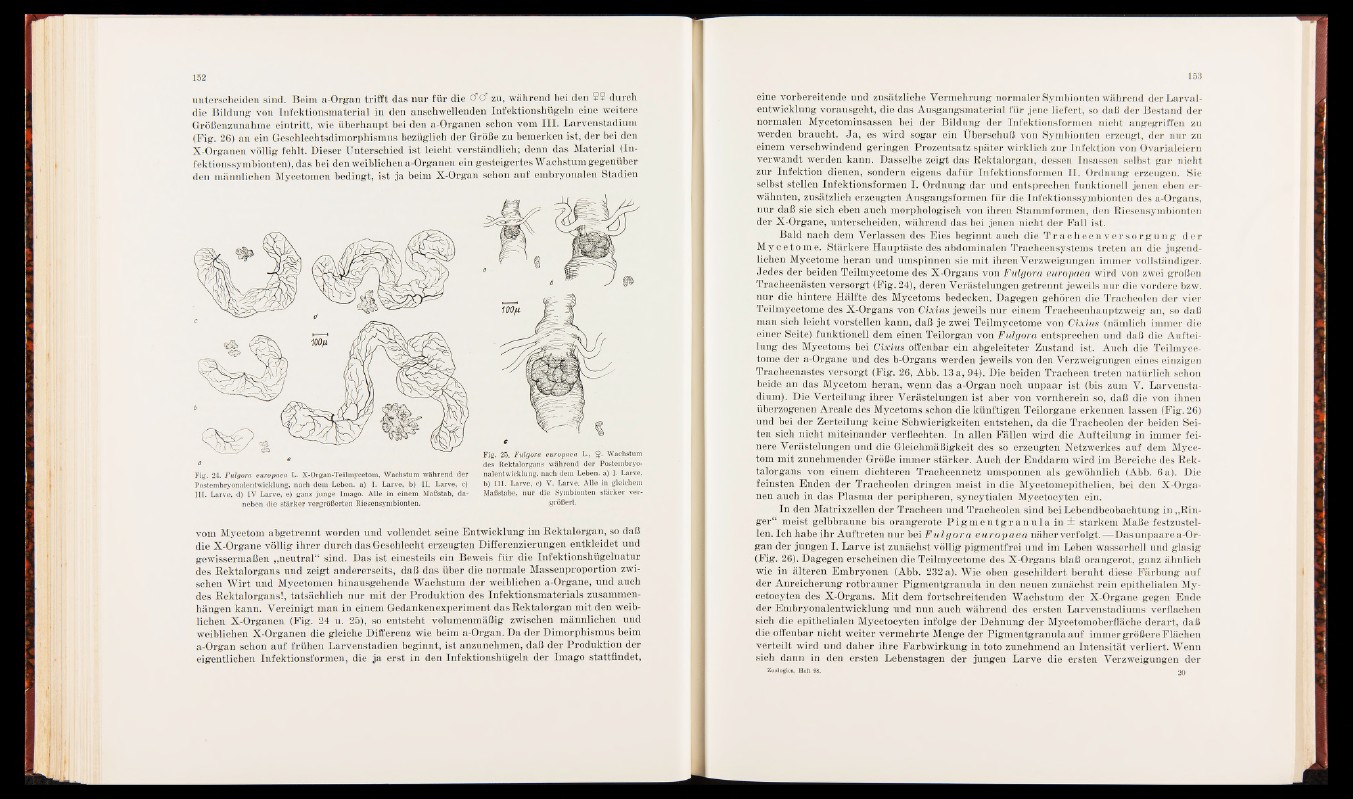
Fig. 24. Fulgora europaea L. X-Organ-Teilmycetom, Wachstum während der
Postembryonalentwicklung, nach dem Leben, a) I. Larve, b) II. Larve, c)
III. Larve, d) IV Larve, e) ganz junge Imago. Alle in einem Maßstab, daneben
die stärker vergrößerten Riesensymbionten.
c
Fig. 25. Fulgora europaea L., $ . Wachstum
des Rektalorgans während der Postembryonalentwicklung,
nach dem Leben, a) I. Larve,
b) III. Larve, c) V. Larve. Alle in gleichem
Maßstabe, nur die Symbionten stärker vergrößert.
vom Mycetom abgetrennt worden und vollendet seine Entwicklung im Rektalorgan, so daß
die X-Organe völlig ihrer durch das Geschlecht erzeugten Differenzierungen entkleidet und
gewissermaßen „neutral“ sind. Das ist einesteils ein Beweis für die Infektionshügelnatur
des Rektalorgans und zeigt andererseits, daß das über die normale Massenproportion zwischen
Wirt und Mycetomen hinausgellende Wachstum der weiblichen a-Organe, und auch
des Rektalorgans!, tatsächlich nur mit der Produktion des Infektionsmaterials Zusammenhängen
kann. Vereinigt man in einem Gedankenexperiment das Rektalorgan mit den weiblichen
X-Organen (Fig. 24 u. 25), so entsteht volumenmäßig zwischen männlichen und
weiblichen X-Organen die gleiche Differenz wie beim a-Organ. Da der Dimorphismus beim
a-Organ schon auf frühen Larvenstadien beginnt, ist anzunehmen, daß der Produktion der
eigentlichen Infektionsformen, die ja erst in den Infektionshügeln der Imago stattfindet,
eine vorbereitende und zusätzliche Vermehrung normaler Symbionten während der Larvalentwicklung
vorausgeht, die das Ausgangsmaterial für jene liefert, so daß der Bestand der
normalen Mycetominsassen bei der Bildung der Infektionsformen nicht angegriffen zu
werden braucht. Ja , es wird sogar ein Überschuß von Symbionten erzeugt, der nur zu
einem verschwindend geringen Prozentsatz später wirklich zur Infektion von Ovarialeiern
verwandt werden kann. Dasselbe zeigt das Rektalorgan, dessen Insassen selbst gar nicht
zur Infektion dienen, sondern eigens dafür Infektionsformen II. Ordnung erzeugen. Sie
seihst stellen Infektionsformen I. Ordnung dar und entsprechen funktionell jenen eben erwähnten,
zusätzlich erzeugten Ausgangsformen für die Infektionssymbionten des a-Organs,
nur daß sie sich eben auch morphologisch von ihren Stammformen, den Riesensymbionten
der X-Organe, unterscheiden, während das bei jenen nicht der Fall ist.
Bald nach dem Verlassen des Eies beginnt auch die T r a c h e e n v e r s o r g u n g de r
My c e t ome . Stärkere Hauptäste des abdominalen Tracheensystems treten an die jugendlichen
Mycetome heran und umspinnen sie mit ihren Verzweigungen immer vollständiger.
Jedes der beiden Teilmycetome des X-Organs von Fulgora europaea wird von zwei großen
Tracheenästen versorgt (Fig. 24), deren Verästelungen getrennt jeweils nur die vordere bzw.
nur die hintere Hälfte des Mycetoms bedecken. Dagegen gehören die Tracheolen der vier
Teilmycetome des X-Organs von Cixius jeweils nur einem Tracheenhauptzweig an, so daß
man sich leicht vorstellen kann, daß je zwei Teilmycetome von Cixius (nämlich immer die
einer Seite) funktionell dem einen Teilorgan von Fulgora entsprechen und daß die Aufteilung
des Mycetoms bei Cixius offenbar ein abgeleiteter Zustand ist. Auch die Teilmycetome
der a-Organe und des b-Organs werden jeweils von den Verzweigungen eines einzigen
Tracheenastes versorgt (Fig. 26, Abb. 13 a, 94). Die beiden Tracheen treten natürlich schon
beide an das Mycetom heran, wenn das a-Organ noch unpaar ist (bis zum V. Larvenstadium).
Die Verteilung ihrer Verästelungen ist aber von vornherein so, daß die von ihnen
überzogenen Areale des Mycetoms schon die künftigen Teilorgane erkennen lassen (Fig. 26)
und hei der Zerteilung keine Schwierigkeiten entstehen, da die Tracheolen der beiden Seiten
sich nicht miteinander verflechten. In allen Fällen wird die Aufteilung in immer feinere
Verästelungen und die Gleichmäßigkeit des so erzeugten Netzwerkes auf dem Mycetom
mit zunehmender Größe immer stärker. Auch der Enddarm w ird im Bereiche des Rektalorgans
von einem dichteren Tracheennetz umsponnen als gewöhnlich (Abb. 6 a). Die
feinsten Enden der Tracheolen dringen meist in die Mycetomepithelien, bei den X-Orga-
nen auch in das Plasma der peripheren, syncytialen Mycetocyten ein.
In den Matrixzellen der Tracheen und Tracheolen sind bei Lebendbeobachtung in „Ringer“
meist gelbbraune bis orangerote P i g m e n t g r a n u l a in ± starkem Maße festzustellen.
Ich habe ihr Auftreten nur bei F u lg o r a e u ro p a e a näher verfolgt.—Das unpaare a-Organ
der jungen I. Larve ist zunächst völlig pigmentfrei und im Leben wasserhell und glasig
(Fig. 26). Dagegen erscheinen die Teilmycetome des X-Organs blaß orangerot, ganz ähnlich
wie in älteren Embryonen (Abb. 232 a). Wie oben geschildert beruht diese Färbung auf
der Anreicherung rotbrauner Pigmentgranula in den neuen zunächst rein epithelialen Mycetocyten
des X-Organs. Mit dem fortschreitenden Wachstum der X-Organe gegen Ende
der Emhryonalentwicklung und nun auch während des ersten Larvenstadiums verflachen
sich die epithelialen Mycetocyten infolge der Dehnung der Mycetomoberfläche derart, daß
die offenbar nicht weiter vermehrte Menge der Pigmentgranula auf immer größere Flächen
verteilt wird und daher ihre Farbwirkung in toto zunehmend an Intensität verliert. Wenn
sich dann in den ersten Lebenstagen der jungen Larve die ersten Verzweigungen der
Zoologien, Heft 98. 2 0