unserer Primärzoide deutlicher markiert, indem das beträchtliche Anwachsen des sexuell determinierten
Larvenabschnitts den „Stolo“ sekundär in einer kreisförmigen Einsenkung der Rücken -
fläche (cf. dieTextfigg. 10, 11 a,b und Fig: 3 (*) Tafel XXXI) unserer Larven erscheinen läßt. Der
Stolostumpf erhält sich nun längere Zeit und erfährt eine Reihe interessanter Umbildungen.
Er verbreitert sich in der Folge beträchtlich (Textfig. 10) und läßt an seinem Scheitelpol
eine drüsige Einsenkung erkennen, die ich in stärkerer Vergrößerung auf Tafel XXXI, Fig. 4 nochmals
abgebildet habe. Die Zellen haben sich aus ihrem Verbände losgelöst und weisen deutliche Zeichen
der Entartung auf. Damit im Zusammenhang scheint die Abgrenzung des fraglichen Bezirks
gegen seine Umgebung durch eine deutliche, strukturlose,
im ungefärbten Präparat schwärzlichbraun erscheinende
Lamelle zu stehen.
Ein Vergleich dieser Lamelle m it der „ chitin“ -artigen Auskleidung
der Gasflasche bei Pneumatophoriden macht es wahrscheinlich,
daß es sich hier wesen tlich um dieselbe Substanz handelt.
Dieser apikalen Drüse kommt in der heutigen
Ontogenese unserer Larven offenbar keine erhebliche
Bedeutung zu. Dagegen würde erstens einmal die Kleinheit
der Drüse sprechen; ferner die fast gleichzeitige
Anlage der als Fallschirm funktionierenden Rückenscheibe
(Schirm, Glocke) am aboralen Pol unserer Primärzoide,
wodurch diese in medusenähnliche Geschöpfe verwandelt
werden.
Daß diese auffällige Isochrome der erwähnten
Vorgänge (Entwicklung einer apikalen Drüse + Anlage
der Rückenscheibe) nicht ein ursprüngliches Verhalten
darstellt, leuchtet wohl ohne weiteres ein. ••
Wenn heutzutage die beschriebene drüsige Einsenkung
keine Rolle in der Ontogenie spielt, so schließt
das keineswegs aus, daß sie früher einmal eine größere
biologische Bedeutung besessen hat.
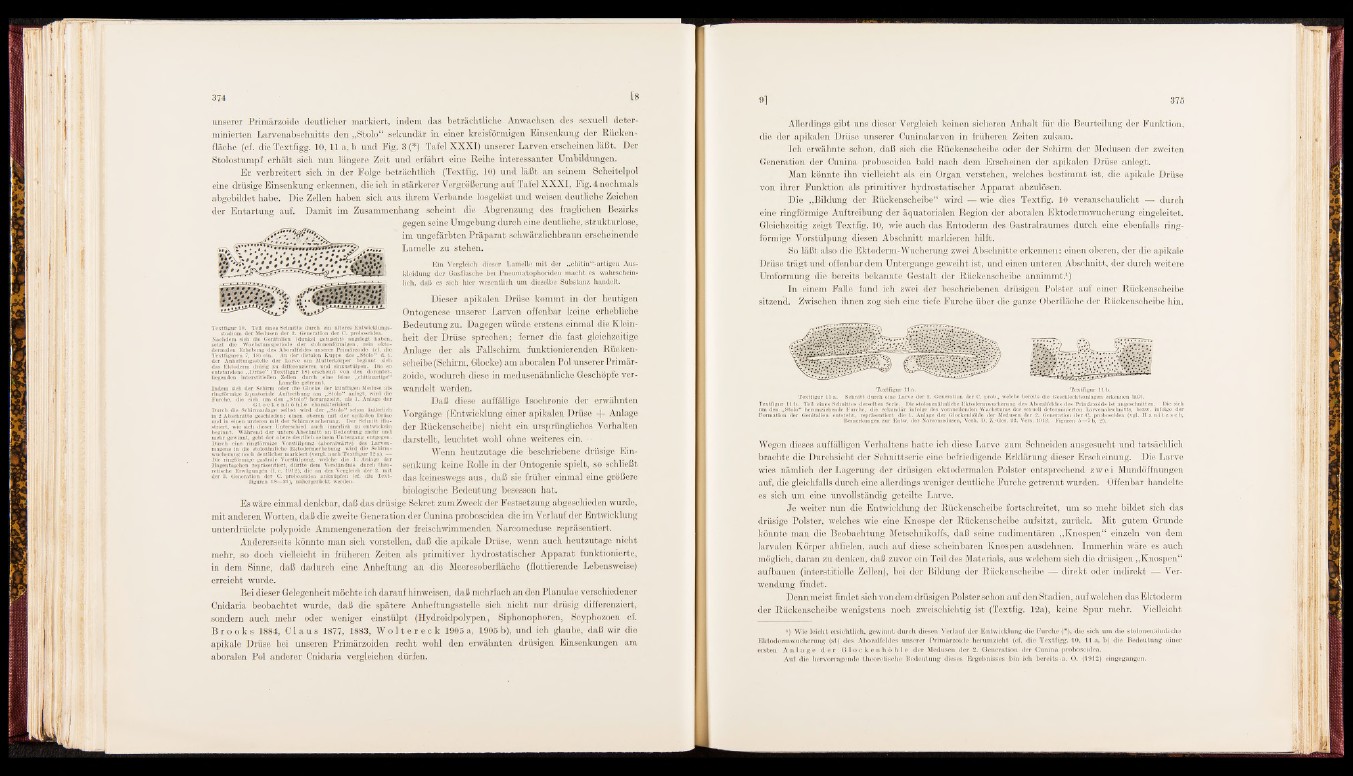
Textfigur 10. Teil eines Schnitts durch ein iUteres Entwicklungsstadium
der Medusen der 2. Generation der C. proboscidea.
Nachdem sich die Genitalien (dunkel getuscht) angelegt haben,
setzt die Wachstumsperiode der stolononförmigen, rein ekto-
dermalen Erhebung des Aboralfeides unserer Primärzoide (cf. die
Textfiguren 7, 18) ein. An der distalen Kuppe des „Stolo“ d. i.
der Anheftungsstelle der Larve am Mutterkörper beginnt sich
das Ektoderm drüsig zu differenzieren und einzustülpen. Die so
entstandene „Drüse“ (Textfigur 18) erscheint von den darunter-
Uegenden interstitiellen Zellen durch eine feine „chitinartige“
Lamelle getrennt.
Indem sich der Schirm oder die Glocke der künftigen Meduse als
ringförmige äquatoriale Auftreibung am „Stolo" anlegt, wird die
Furche, die sich um den „Stolo^herumzieht, als 1. Anlage der
G lo c k e n.n öIi-Lc charakterisiert.
Durch die Schinnanlage selbst wird der „Stolo“ schon äußerlich
in 2 Abschnitte geschieden; einen oberen mit der apikalen Drüse
und in einen unteren mit der Schirmwucherung. Der Schnitt illustriert,
wie sich dieser Unterschied auch innerlich zu entwickeln
beginnt. Während der untere Abschnitt an Bedeutung mehr und
mehr gewinnt, geht der obere deutlich seinem Untergang entgegen.
Durch eine ringförmige Vorstülpung (aboralwärts) des Laryenmagens
in die stoloähnliche Ektodermerhebung wird die Schirmwucherung
noch deutlicher markiert (vergl. auch Textfigur 12 a). —
Die ringförmige gastrale Vorstülpung, welche die 1. Anlage der
Magentaschen repräsentiert, dürfte dem Verständnis durch theoretische
Erwägungen (1. c. 1912), die an den Vergleich der 2. mit
der 8. Generation der C. proboscidea anknüpfen (cf. die Textfiguren
18—21), näliergerückt werden.
Es wäre einmal denkbar, daß das drüsige Sekret zum Zweck der Festsetzung abgeschieden wurde,
mit anderen Worten, daß die zweite Generation der Cunina proboscidea die im Verlauf der Entwicklung
unterdrückte polypoide Ammengeneration der freischwimmenden Narcomeduse repräsentiert.
Andererseits könnte man sich vorstellen, daß die apikale Drüse, wenn auch heutzutage nicht
mehr, so doch vielleicht in früheren Zeiten als primitiver hydrostatischer Apparat funktionierte,
in dem Sinne, daß dadurch eine Anheftung an die Meeresoberfläche (flottierende Lebensweise)
erreicht wurde.
Bei dieser Gelegenheit möchte ich darauf hinweisen, daß mehrfach an den Planulae verschiedener
Cnidaria beobachtet wurde, daß die spätere Anheftungsstelle sich nicht nur drüsig differenziert,
sondern auch mehr oder weniger einstülpt (Hydroidpolypen, Siphonophoren, Scyphozoen cf.
Br o o k s 1884, Cl aus 1877, 1883, Wo l t e r e c k 1905 a, 1905 b), und ich glaube, daß wir die
apikale Drüse bei unseren Primärzoiden recht wohl den erwähnten drüsigen Einsenkungen am
aboralen Pol anderer Cnidaria vergleichen dürfen.
Allerdings gibt uns dieser Vergleich keinen sicheren Anhalt für die Beurteilung der Funktion,
die der apikalen Drüse unserer Cuninalarven in früheren Zeiten zukam.
Ich erwähnte schon, daß sich die Rückenscheibe oder der Schirm der Medusen der zweiten
Generation der Cunina proboscidea bald nach dem Erscheinen der apikalen Drüse anlegt.
Man könnte ihn vielleicht als ein Organ verstehen, welches bestimmt ist, die apikale Drüse
von ihrer Funktion als primitiver hydrostatischer Apparat abzulösen.
Die „Bildung der Rückenscheibe“ wird — wie dies Textfig. 10 veranschaulicht -— durch
eine ringförmige Auftreibung der äquatorialen Region der aboralen Ektodermwucherung eingeleitet.
Gleichzeitig zeigt Textfig. 10, wie auch das Entoderm des Gastralraumes durch eine ebenfalls ringförmige
Vorstülpung diesen Abschnitt markieren hilft.
So läßt also die Ektoderm-Wucherung zwei Abschnitte erkennen: einen oberen, der die apikale
Drüse trägt und offenbar dem Untergange geweiht ist, und einen unteren Abschnitt, der durch weitere
Umformung die bereits bekannte Gestalt der Rückenscheibe annimmt.1)
In einem Falle fand ich zwei der beschriebenen drüsigen Polster auf einer Rückenscheibe
sitzend. Zwischen ihnen zog sich eine tiefe Furche über die ganze Oberfläche der Rückenscheibe hin.
Bemerkungen z
i der G. prob., welche bereits die Gesell
mähnliche Ektodermwucherung des Aboralfeldes d
| s der Glockenhöhle der Medusen der 2. Generatio
der Narcomodusen, Verh. D. Z.-Ges. 22. Vers. 1912. Figif
Wegen dieses auffälligen Verhaltens hatte ich diese Larve zum Schneiden ausgesucht und tatsächlich
brachte die Durchsicht der Schnittserie eine befriedigende Erklärung dieser Erscheinung. Die Larve
wies nämlich der Lagerung der drüsigen ektodermalen Polster entsprechend zw e i Mundöffnungen
auf, die gleichfalls durch eine allerdings weniger deutliche Furche getrennt wurden. Offenbar handelte
es sich um eine unvollständig geteilte Larve.
Je weiter nun die Entwicklung der Rückenscheibe fortschreitet, um so mehr bildet sich das
drüsige Polster, welches wie eine Knospe der Rückenscheibe aufsitzt, zurück. Mit gutem Grunde
könnte man die Beobachtung Metschnikoffs, daß seine rudimentären „Knospen“ einzeln von dem
larvalen Körper abfielen, auch auf diese scheinbaren Knospen ausdehnen. Immerhin wäre es auch
möglich, daran zu denken, daß zuvor ein Teil des Materials, aus welchem sich die drüsigen „Knospen“
auf bauen (interstitielle Zellen), bei der Bildung der Rückenscheibe — direkt oder indirekt — Verwendung
findet.
Denn meist findet sich von dem drüsigen Polster schon auf den Stadien, auf welchen das Ektoderm
der Rückenscheibe wenigstens noch zweischichtig ist (Textfig. 12a), keine Spur mehr. Vielleicht
*) Wie leich t ersichtlich, gewinnt durch diesen Verlauf der Entwicklung die Furche (*), die sich um die stolonenähnliche
Ektodermwucherung (st) des Aboralfeldes unserer Primärzoide herumzieht (cf. die Textfigg. 10, 1 1 a , b) die Bedeutung einer
ersten A n l a g e d e r G l o c k e n h ö h l e der Medusen der 2. Generation der Cunina proboscidea.
Auf die hervorragende theoretische Bedeutung dieses Ergebnisses bin ich bereits -a. .0. (1912) eingegangen.