durch einen Zeitraum hindurch in Ruhe verharren, der die Gesamtdauer der voraus-
gegangenen Klüftung um ein vielfaches übertrifft?
V o r allem aber ist ja das Soma mit seinen einzelnen ungleichwertigen Zellfamilien
der eigentliche Schauplatz der rhythmischen Differenzierung. Und hier im Soma sind alle
Kerne, so viel man in den frühen Stadien erkennen kann, untereinander gleich. Die Art
ihrer Umwandlung, der Bau ihrer endlichen Reifezustände und, wofern es sich um dieselbe
Generationsstufe handelt, auch ihre maximale Größe stimmen überein. Nur eben die G e s
c h w in d i g k e i t ihres Herausreifens ist typisch verschieden. Und es gibt keinen markanteren
Fall, das ungleiche Reifetempo gleich beschaffener Kerne zu demonstrieren, als die
schon vorhin erwähnte rhythmische Divergenz der Urzellen des Schlundes und des primären
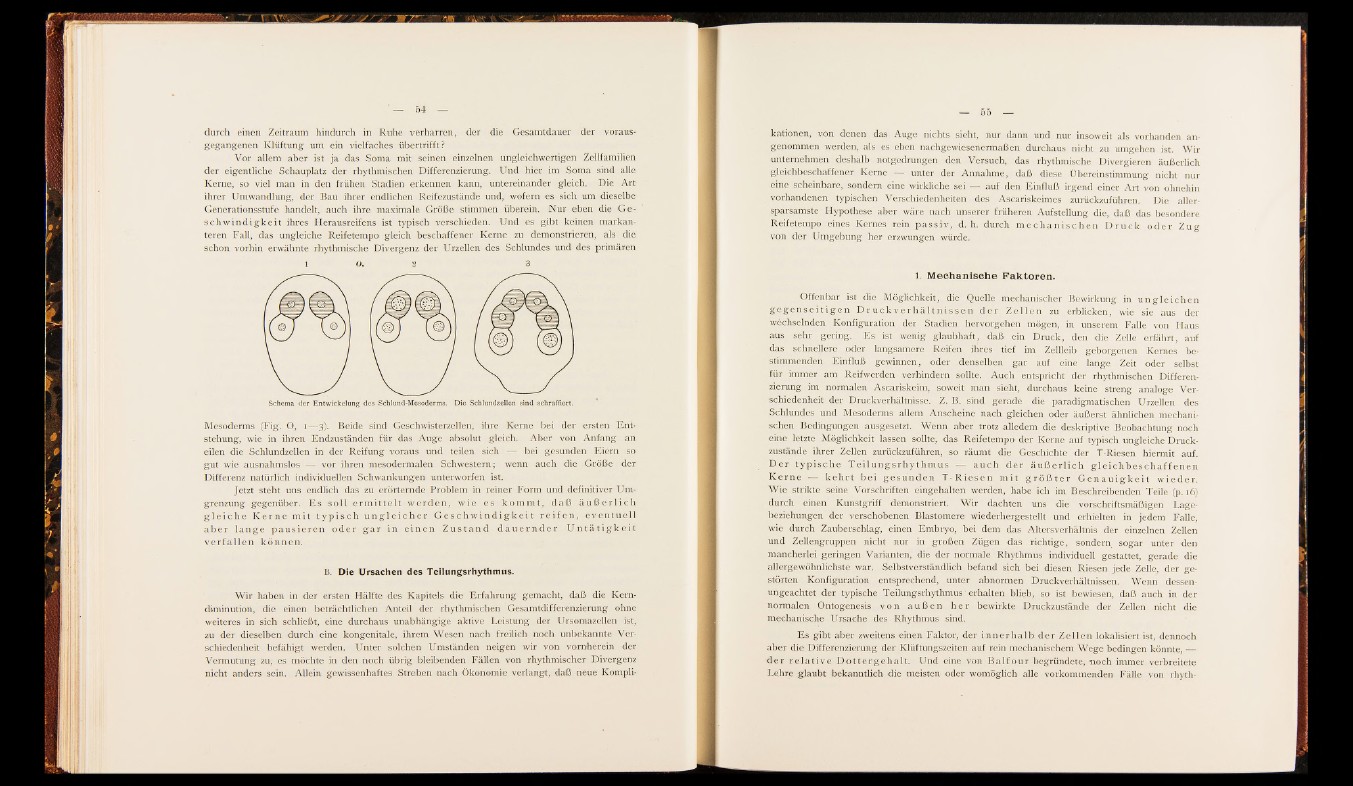
Schema der Entwickelung des Schlund-Mesoderms. Die Schlundzellen sind schraffiert.
Mesoderms (Fig. O, i— -3). Beide sind Geschwisterzellen, ihre Kerne bei der ersten E n tstehung,
wie in ihren Endzuständen für das Ä u g e absolut gleich. Aber von A nfan g an
eilen die Schlundzellen in der Reifung voraus und teilen sich — bei gesunden Eiern so
gut wie ausnahmslos -|f| vor ihren mesodermalen Schwestern; wenn auch die Größe der
Differenz natürlich individuellen Schwankungen unterworfen ist.
Jetzt steht uns endlich das zu erörternde Problem in reiner Form und definitiver Umgrenzung
gegenüber. E s s o l l e rm i t t e l t w e r d e n , w i e e s k o m m t , d a ß ä u ß e r l i c h
g l e i c h e K e r n e m i t t y p i s c h u n g l e i c h e r G e s c h w i n d i g k e i t r e i f e n , e v e n t u e l l
a b e r l a n g e p a u s i e r e n o d e r g a r in e in e n Z u s t a n d d a u e r n d e r U n t ä t i g k e i t
v e r f a l l e n k ö n n e n .
B. Die Ursachen des Teilungsrhythmus.
W ir haben in der ersten Hälfte des Kapitels die Erfahrung gemacht, daß die Kern-
diminution, die einen beträchtlichen Anteil der rhythmischen Gesamtdifferenzierung ohne
weiteres in sich schließt, eine durchaus unabhängige aktive Leistung der Ursomazellen ist,
zu der dieselben durch eine kongenitale, ihrem Wesen nach freilich noch unbekannte V e r schiedenheit
befähigt werden. Unter solchen Umständen neigen wir von vornherein der
Vermutung zu, es möchte in den noch übrig bleibenden Fällen von rhythmischer Divergenz
nicht anders sein. Allein gewissenhaftes Streben nach Ökonomie verlangt, daß neue Komplikationen,
von denen das A uge nichts sieht, nur dann und nur insoweit als vorhanden angenommen
werden, als es eben nachgewiesenermaßen durchaus nicht zu umgehen ist. Wir
unternehmen deshalb notgedrungen den Versuch, das rhythmische Divergieren äußerlich
gleichbeschaffener Kerne — unter der Annahme, daß diese Übereinstimmung nicht nur
eine scheinbare, sondern eine wirkliche sei — auf den Einfluß irgend einer Art von ohnehin
vorhandenen typischen Verschiedenheiten des Ascariskeimes zurückzuführen. Die allersparsamste
Hypothese aber wäre nach unserer früheren Aufstellung die, daß das besondere
Reifetempo eines Kernes rein p a s s i v , d .h . durch m e c h a n i s c h e n D r u c k o d e r Z u g
von der Umgebung her erzwungen würde.
l. M e c h a n is c h e F a k to r e n .
Offenbar ist die'¿Möglichkeit, die Quelle mechanischer Bewirkung in u n g l e i c h e n
g e g e n s e i t i g e n D r u c k v e r h ä l t n i s s e n d e r Z e l l e n zu erblicken, wie sie aus der
wechselnden Konfiguration der Stadien hervorgehen mögen, in unserem Falle von Haus
aus sehr gering. E s ist wenig glaubhaft, daß ein Druck, den die Zelle erfährt, auf
das schnellere oder langsamere Reifen ihres tief im Zellleib geborgenen Kernes bestimmenden
Einfluß gewinnen, oder denselben gar auf eine lange Zeit oder selbst
für immer am Reifwerden verhindern sollte. A uch entspricht der rhythmischen Differenzierung
im normalen Ascariskeim, soweit man sieht, durchaus keine streng analoge Verschiedenheit
der Druckverhältnisse. Z. B. sind gerade die paradigmatischen Urzellen des
Schlundes und Mesoderms allem Anscheine nach gleichen oder äußerst ähnlichen mechanischen
Bedingungen ausgesetzt. Wenn aber trotz alledem die deskriptive Beobachtung noch
eine letzte Möglichkeit lassen sollte, das Reifetempo der Kerne auf typisch ungleiche Druckzustände
ihrer Zellen zurückzuführen, so räumt die Geschichte der T-Riesen hiermit auf.
D e r t y p i s c h e T e i l u n g s r h y t h m u s — a u c h d e r ä u ß e r l i c h g l e i c h b e 's c h a f f e n e n
K e r n e g k e h r t b e i g e s u n d e n T - R i e s e n m i t g r ö ß t e r G e n a u i g k e i t w i e d e r .
W ie strikte seine Vorschriften ein'gehalten werden, habe ich im Beschreibenden Teile (p. 16)
durch einen Kunstgriff demonstriert. Wir dachten uns die vorschriftsmäßigen La g e beziehungen
der verschobenen Blastomere Wiederhergestellt und erhielten in jedem Falle,
wie durch Zauberschlag, einen Embryo, bei dem das Altersverhältnis der einzelnen Zellen
und Zellengruppen nicht nur in großen Zügen das richtige, sondern sogar unter den
mancherlei geringen Varianten, die der normale Rhythmus individuell gestattet, gerade die
allergewöhnlichste war. Selbstverständlich befand sich bei diesen Riesen jede Zelle, der ge störten
Konfiguration entsprechend, unter abnormen Druckverhältnissen. Wenn dessenungeachtet
der typische Teilungsrhythmus’ erhalten blieb, so ist bewiesen, daß auch in der
normalen Ontogenesis v o n a u ß e n h e r bewirkte Druckzustände der Zellen nicht die
mechanische Ursache des Rhythmus sind.
E s gibt aber zweitens einen Faktor, der in n e r h a lb d e r Z e l l e n lokalisiert ist, dennoch
aber die Differenzierung der Klüftungszeiten auf rein mechanischem W e g e bedingen könnte,
d e r r e l a t i v e D o t t e r g e h a l t . Und eine von B a l f o u r begründete, no ch immer verbreitete
Lehre glaubt bekanntlich die meisten oder womöglich alle vorkommenden Fälle von rhyth