von einigen F o r s ch em für „ s y m p a t i s c h “ gehalten und findet sich auch bei manchen anderen
Arachniden. Zwei zarte Nervenpaare innervieren endlich die C o x a ld r ü s e n (bei Thelyphoniden
und Tarantuliden)^ sie entspringen auf der dorsalen Fläche des Unterschlundganglions, ihre
Segmentzugehörigkeit ist aber noch nicht ermittelt worden.
Das O p i s t h o s o m a enthält entweder noch ein eigenes Ganglion oder es entbehrt derselben
vollständig. Sein höchstens in der Einzahl vorhandenes Ganglion liegt bei Thelyphoniden
etwa an der Grenze des 8. und 9. Segmentes, bei Schizonotiden und Koenenien im Genitalsegment,
und ist bei den beiden erstgenannten Formen durch eine Kommissur, bei Koenenia unmittelbar
mit dem prosomalen Untersehlundganglion verbunden. Die Tarantuliden haben kein opis-
thosomales Ganglion mehr, und sie stellen zweifellos bezüglich des Nervensystems die am
meisten abgeleiteten, resp. im Sinne der Konzentration der Ganglien höchstentwickelten Pedi-
palpen dar, indem sie gleichzeitig zu den echten Araneen überleiten. Trithyreus und Koenenia
vermitteln ihrerseits zwischen Thelyphonen und Tarantuliden. & |||||
Die bis heute allgemein herrschende Ansicht, daß The/yphonus infolge des Besitzes eines
Hinterleibsganglions im Bau des Nervensystems u r s p ü n g l i c h e r s e i a l s die Tarantuliden,
hat nun in jüngster Zeit R. J. P o c o c k (53) fallen lassen. E r stützt sich dabei auf die verschiedenartige
Innervierung der vorderen und hinteren Segmente des Hinterleibes und sagt,
daß „in the Thelyphonidae it seems clear that the innervation o f the flexible posterior end
o f the opisthosoma is the sole function o f the median cord. If these organs were suppressed,
the nervecord (Hinterleibsganglion samt seiner Kommissur) would, become useless and might
cease to be developped. The whole o f the sternal surface o f the opisthosoma would then
receive its nervous supply from the cords I have above described, (den Seitennerven), which
would certainly be taken for the primitive median co rd, although they would in reality represent
merely its original laterally and metamerically diverging threads.“
E s ist nicht schwer, diese Auffassung zu widerlegen, da sie lediglich auf einem Mißverständnis
der beiden seitlichen Nervenstränge beruht, welche bei Thelyphoniden und Tarantuliden
die vorderen Hinterleibssegmente innervieren. Um diese richtig zu verstehen, gehen
wir am besten von dem denkbar einfachsten Verhältnis des opisthosomalen Nervensystems,
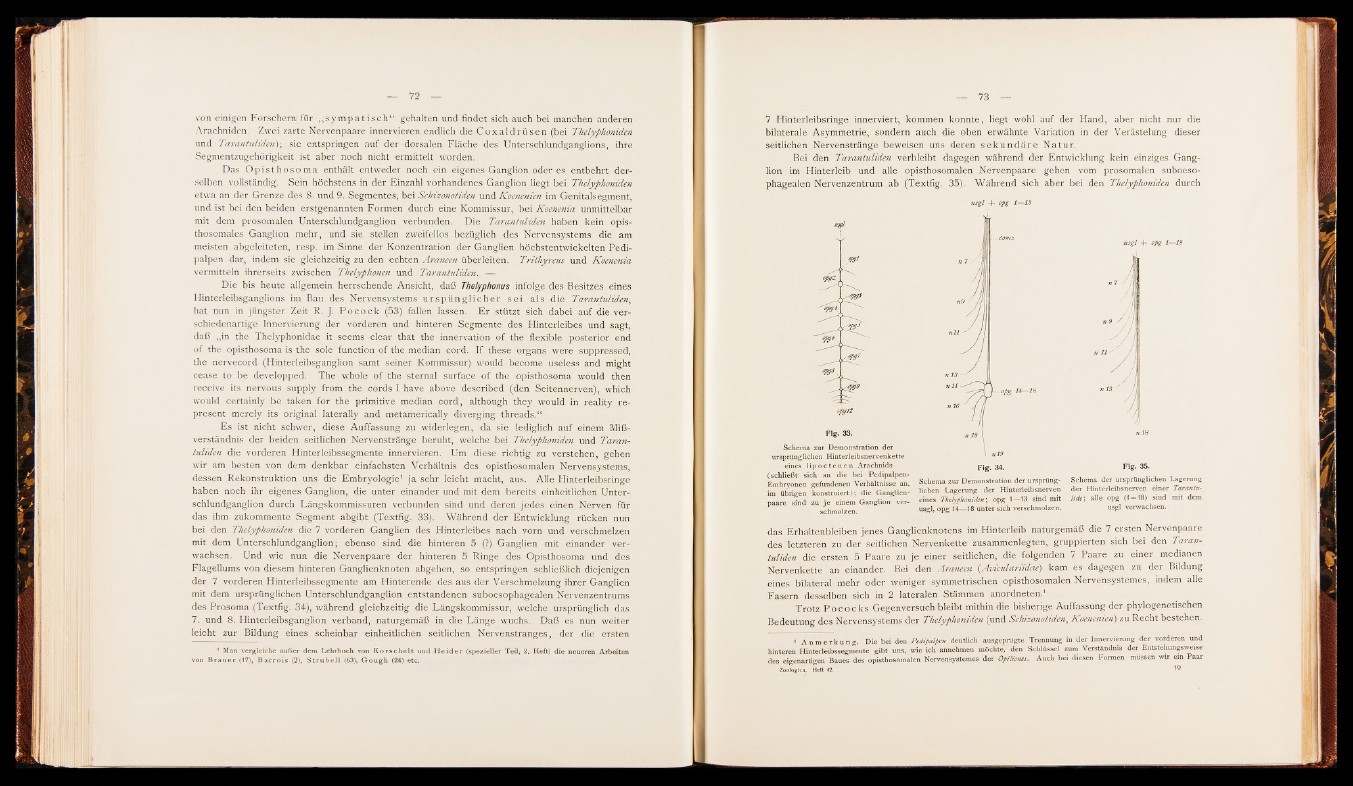
dessen Rekonstruktion uns die Embryologie1 ja sehr leicht macht, aus. A lle Hinterleibsringe
haben noch ihr eigenes Ganglion, die unter einander und mit dem bereits einheitlichen Unterschlundganglion
durch Längskommissuren verbunden sind und deren jedes einen Nerven für
das ihm zukommente Segment abgibt (Textfig. 33). Während der Entwicklung rücken nun
bei den Thelyphoniden die 7 vorderen Ganglien des Hinterleibes nach vorn und verschmelzen
mit dem Untersehlundganglion; ebenso sind die hinteren 5 (?) Ganglien mit einander verwachsen.
Und wie nun die Nervenpaare der hinteren 5 Ringe des Opisthosoma und des
Flagellums von diesem hinteren Ganglienknoten abgehen, so entspringen schließlich diejenigen
der 7 vorderen Hinterleibssegmente am Hinterende des aus der Verschmelzung ihrer Ganglien
mit dem ursprünglichen Untersehlundganglion entstandenen suboesophagealen Nervenzentrums
des Prosoma (Textfig. 34), während gleichzeitig die Längskommissur, welche ursprünglich das
7. und 8. Hinterleibsganglion verband, naturgemäß in die Län ge wuchs. Daß es nun weiter
leicht zur Bildung eines scheinbar einheitlichen seitlichen Nervenstranges, der die ersten
1 Man vergleiche außer dem Lehrbuch von K o r s c h e lt und H e id e r (spezieller Teil, 2. Heft) die neueren Arbeiten
von B r a u e r (17), B a r r o is (2), S t ru b e i l (63), G o u g h (24) etc.
7 Hinterleibsringe innerviert, kommen konnte.*; liegt wohl auf der Hand, aber nicht nur die
bilaterale Asymmetrie, sondern auch die oben erwähnte Variation in der Verästelung dieser
seitlichen Nervenstränge beweisen uns deren s e k u n d ä r e N a tu r .
Bei den Tarantuliden verbleibt dagegen während der Entwicklung kein einziges Ganglion
im Hinterleib und alle opisthosomalen Nervenpaare gehen vom prosomalen suboesophagealen
Nervenzentrum ab (Textfig. 35). Während sich aber bei den Thelyphoniden durch
usgl + opg 1—13
usgl + Opg 1—18
opglZ
Flg. 33.
Schema zur Demonstration der
ursprünglichen Hinterleibsnervenkette
eines l ip o c t e n e n Arachnids
(schließt sich an die bei Pedipalpen-
Fig. 34. Flg. 35.
, T
imbryonen gefundenen Verhältnisse an, Schema B Demonstration der Ursprung- Schema der ursprünglichen Lagerung
im übrigen konstruiert); die Ganglien- Sehen Lagerung der Hinterleibsnerven der Hmterleibsnerven einer Taranto-
paare sind zu je einem Ganglion ver- eines ThelyfhMidM-, opg 1 - 1 3 sind mit * * ; alle opg ( 1 - 18 ) sind mit dem
schmolzen. usgl, opg 14— 18 unter sich verschmolzen. usgl verwachsen.
das Erhaltenbleiben jenes Ganglienknotens im Hinterleib naturgemäß die 7 ersten Nervenpaare
des letzteren zu der seitlichen Nervenkette zusammenlegten, gruppierten sich bei den Tarantuliden
die ersten 5 Paare zu je einer seitlichen, die folgenden 7 Paare zu einer medianen
Nervenkette an einander. Bei den Araneen (Aviculanidae) kam es dagegen zu der Bildung
eines bilateral mehr oder weniger symmetrischen opisthosomalen Nervensystemes, indem alle
Fasern desselben sich in 2 lateralen Stämmen anordneten.1
Trotz P o c o c k s Gegenversuch bleibt mithin die bisherige Auffassung der phylogenetischen
Bedeutung des Nervensystems der Thelyphoniden (und Schizonotiden, Koenenien) zu Recht bestehen.
i A n m e r k u n g . Die bei den Pedipalpen deutlich ausgeprägte Trennung in der Innervierung der vorderen und
hinteren Hinterleibssegmente gibt uns, wie ich annehmen möchte, den Schlüssel zum Verständnis der Entstehungsweise
des eigenartigen Baues des opisthosomalen Nervensystemes der Opiliones. Auch bei diesen Formen müssen ’
Zoologica. Heft 42.
wir ein Paar